Eicosanoids are a structurally diverse family of lipid mediators derived from C20 polyunsaturated fatty acids — primarily arachidonic acid (AA, 20:4n-6), eicosapentaenoic acid (EPA, 20:5n-3), and dihomo-γ-linolenic acid (DGLA, 20:3n-6). These potent signaling molecules operate locally at nanomolar to picomolar concentrations, regulating inflammation, vascular tone, platelet aggregation, and immune cell trafficking through G protein-coupled receptors expressed on target cells. Their biosynthesis is not constitutive but is triggered by cell-specific stimuli — inflammatory cytokines, growth factors, and mechanical stress — that activate phospholipase A2 enzymes to release AA from membrane phospholipids. Once synthesized, eicosanoids act locally, are rapidly metabolized to inactive products, and are excreted in urine as stable metabolites. The existing resource on this topic provides a comprehensive survey of eicosanoid classes, biosynthetic pathways, metabolic routes, disease associations, and mass spectrometry techniques. This rewritten guide preserves and extends that content by integrating the descriptive biology of eicosanoid classes and functions with the quantitative analytical framework required to design and interpret eicosanoid profiling experiments, covering sample collection workflows, LC-MS/MS method optimization, lipid mediator class switching over the time course of inflammation, and pharmacodynamic biomarker strategies for preclinical drug discovery.
Eicosanoid analysis services provide validated LC-MS/MS methods for quantitative profiling of prostaglandins, leukotrienes, thromboxanes, lipoxins, and specialized pro-resolving mediators.
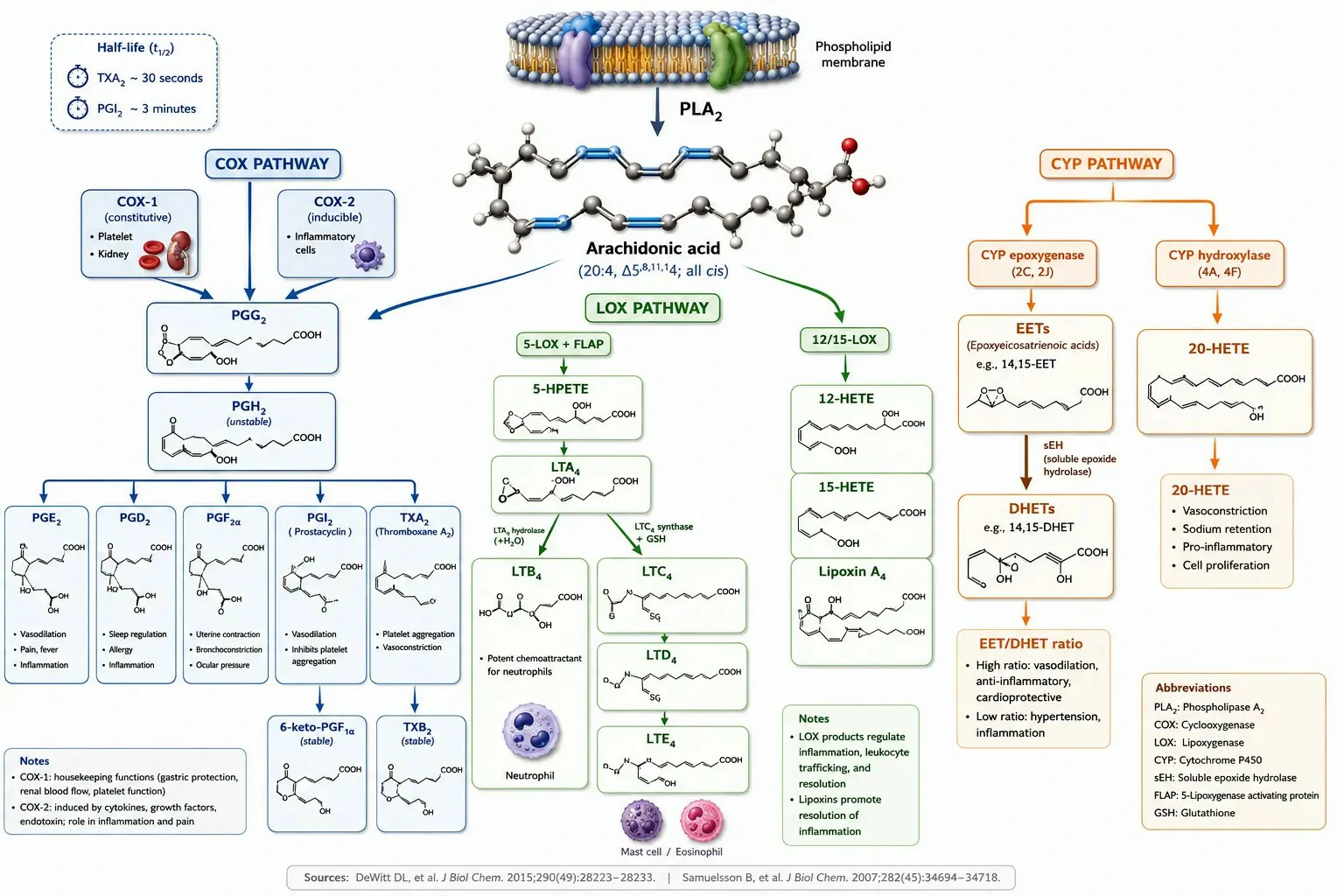
Figure 1: Eicosanoid biosynthesis overview — COX, LOX, and CYP pathways with key enzyme isoforms, major products, and stable metabolites
What Are Eicosanoids and Their Major Classes
Eicosanoids are oxygenated derivatives of C20 polyunsaturated fatty acids that function as local hormones — they are synthesized on demand, act on nearby target cells through specific receptors, and are rapidly inactivated rather than circulating as stable endocrine signals. The term "eicosanoid" derives from the Greek "eikosi" (twenty), referring to the 20-carbon fatty acid precursors from which they are synthesized.
Prostaglandins (PGs) are the most diverse eicosanoid class, all sharing a 20-carbon skeleton that contains a cyclopentane ring formed by the cyclooxygenase reaction. The major prostaglandins are distinguished by the substituents on the cyclopentane ring: PGE2 (a 9-keto, 11,15-dihydroxy structure) mediates inflammation, pain sensitization, and fever through EP receptors; PGD2 (a 9,15-dihydroxy, 11-keto structure) is the major prostaglandin produced by mast cells and mediates allergic responses and sleep regulation through DP receptors; PGF2α (9,11,15-trihydroxy) stimulates uterine contraction and luteolysis through FP receptors; and PGI2 (prostacyclin, containing a second ring formed by the 6,9-epoxy group) is a potent vasodilator and inhibitor of platelet aggregation produced by vascular endothelial cells, acting through IP receptors. Prostaglandins are chemically unstable — PGI2 has a half-life of approximately 3 minutes in aqueous solution at physiological pH, hydrolyzing non-enzymatically to the stable but biologically inactive 6-keto-PGF1α.
Thromboxanes (TXs) are structurally related to prostaglandins but contain a six-membered oxane ring instead of the five-membered cyclopentane ring. TXA2, the major thromboxane, is produced primarily by platelets and is the most potent known inducer of platelet aggregation and vasoconstriction, acting through TP receptors. TXA2 is the most unstable of all eicosanoids, with a half-life of approximately 30 seconds in aqueous solution, hydrolyzing non-enzymatically to the stable TXB2. This extreme instability means that TXA2 acts strictly locally — on platelets within the same thrombus — and that all analytical measurements of thromboxane production rely on quantifying its stable metabolite TXB2 or its urinary metabolite 11-dehydro-TXB2.
Leukotrienes (LTs) are produced by the 5-lipoxygenase pathway and contain a conjugated triene chromophore that gives the class its name. LTB4 (5,12-dihydroxy-eicosatetraenoic acid) is a potent chemoattractant and activator of neutrophils, produced by neutrophils and macrophages and acting through BLT1 and BLT2 receptors. The cysteinyl leukotrienes — LTC4, LTD4, and LTE4 — contain a glutathione, cysteinylglycine, or cysteine group conjugated to the eicosanoid backbone at position 6 and are produced predominantly by mast cells, eosinophils, and basophils. They are the major mediators of bronchoconstriction in asthma, acting through CysLT1 and CysLT2 receptors to cause airway smooth muscle contraction, increased vascular permeability, and mucus secretion. LTE4 is the most stable cysteinyl leukotriene and is excreted in urine, where it serves as a biomarker of systemic cysteinyl leukotriene production.
Lipoxins (LXs) are trihydroxy-tetraene eicosanoids that function as stop signals for inflammation. LXA4 and LXB4 are produced through transcellular biosynthesis — 15-LOX in epithelial cells or monocytes generates 15-HETE from arachidonic acid, which is then converted to lipoxins by 5-LOX in neutrophils recruited to the site. Alternatively, platelet 12-LOX can convert neutrophil-derived LTA4 to lipoxins through a second transcellular route. Lipoxins inhibit neutrophil chemotaxis and adhesion, promote neutrophil apoptosis, and enhance macrophage phagocytosis of apoptotic neutrophils (efferocytosis), actively terminating the inflammatory response rather than simply allowing it to decay passively.
Specialized pro-resolving mediators (SPMs) include resolvins, protectins, and maresins, which are produced from EPA and DHA rather than from arachidonic acid. Resolvin D1 (from DHA) and Resolvin E1 (from EPA) are among the most potent pro-resolving mediators, active at concentrations as low as 10-100 pg/mL. They promote the clearance of neutrophils from inflamed tissues, enhance macrophage efferocytosis, and stimulate tissue regeneration. Protectins are DHA-derived mediators characterized by a conjugated triene system and a 10,17-dihydroxy structure; protectin D1 (also known as neuroprotectin D1 when produced in neural tissue) is anti-inflammatory and neuroprotective. Maresins (macrophage mediators in resolving inflammation) are produced by macrophages from DHA through 12-LOX activity and promote tissue regeneration and pain resolution.
Hydroxy fatty acids (HETEs) are produced by lipoxygenase and cytochrome P450 enzymes through the insertion of a single hydroxyl group at various positions on the arachidonic acid chain. 5-HETE, 12-HETE, and 15-HETE are produced by the corresponding LOX enzymes and serve as both signaling molecules in their own right and as intermediates for further enzymatic conversion to leukotrienes and lipoxins. 20-HETE, produced by CYP4A and CYP4F enzymes, is a potent vasoconstrictor that regulates renal and cerebral blood flow and is implicated in hypertension. HETEs are more stable than prostaglandins and leukotrienes and are useful as pathway markers — an increase in 5-HETE without a corresponding increase in LTB4 indicates 5-LOX activation with efficient downstream conversion, while a parallel increase in both suggests a defect in LTA4 hydrolase activity.
Biological Functions of Eicosanoids
Eicosanoids exert their biological effects by binding to specific G protein-coupled receptors on target cells, with each eicosanoid having one or more dedicated receptors that determine its functional profile. The distribution of these receptors across tissues explains the diverse and sometimes opposing effects of different eicosanoids.
Inflammation and immune response: PGE2 is the dominant pro-inflammatory prostaglandin, causing vasodilation and increased vascular permeability that produce the redness, heat, and swelling characteristic of acute inflammation. PGE2 also sensitizes peripheral nociceptors to bradykinin and other pain mediators, contributing to inflammatory hyperalgesia. LTB4 is the primary leukocyte chemoattractant among the eicosanoids, directing neutrophil migration to sites of infection or tissue injury along a concentration gradient. PGD2, produced by mast cells activated by IgE cross-linking, contributes to allergic inflammation. In the resolution phase of inflammation, lipoxins and resolvins actively suppress further neutrophil recruitment and promote the clearance of apoptotic cells, preventing the transition from acute to chronic inflammation.
Vascular tone and blood flow regulation: Eicosanoids exert opposing effects on vascular smooth muscle that allow for regional blood flow regulation. PGI2 produced by endothelial cells is a potent vasodilator that increases local blood flow. PGE2 is also vasodilatory in most vascular beds. TXA2 produced by activated platelets is a potent vasoconstrictor that reduces blood flow at sites of vascular injury, contributing to hemostasis. The cysteinyl leukotrienes constrict vascular smooth muscle and increase vascular permeability, contributing to the edema characteristic of acute inflammation. EETs produced by endothelial CYP epoxygenases are vasodilatory and are considered endothelium-derived hyperpolarizing factors (EDHFs) that contribute to flow-mediated vasodilation independent of nitric oxide and prostacyclin.
Platelet aggregation and hemostasis: The balance between TXA2 (pro-aggregatory, produced by platelets) and PGI2 (anti-aggregatory, produced by endothelial cells) determines the thrombotic tendency at the vessel wall. TXA2 amplifies platelet activation by binding to TP receptors on the platelet surface, triggering shape change, granule secretion, and additional TXA2 synthesis in a positive feedback loop. PGI2 inhibits platelet aggregation by activating platelet IP receptors, which couple to Gs proteins and increase intracellular cAMP. Low-dose aspirin irreversibly inhibits platelet COX-1, blocking TXA2 production for the lifespan of the platelet (7-10 days), while endothelial cells, which can synthesize new COX-1 and COX-2 protein, recover PGI2 production within hours.
Fever and pain sensitization: PGE2 acts on EP3 receptors in the preoptic area of the hypothalamus to raise the thermoregulatory set point, producing fever. This is a centrally mediated effect — PGE2 produced at peripheral inflammatory sites does not cross the blood-brain barrier, but circulating cytokines (IL-1β, IL-6) induce COX-2 expression in brain endothelial cells, which produce PGE2 that acts on hypothalamic neurons. PGE2 also sensitizes peripheral nociceptors by reducing the threshold for action potential generation — an effect that converts normally innocuous stimuli (such as gentle touch or mild temperature changes) into painful stimuli, contributing to the hyperalgesia of inflamed tissues.
Bronchoconstriction and airway inflammation: The cysteinyl leukotrienes LTC4, LTD4, and LTE4 are approximately 1,000-fold more potent than histamine as bronchoconstrictors, making them the major mediators of allergen-induced airway narrowing in asthma. They are produced by mast cells (LTC4), eosinophils, and basophils upon allergen challenge and act on CysLT1 receptors on airway smooth muscle to cause sustained bronchoconstriction that is not relieved by antihistamines but is blocked by leukotriene receptor antagonists such as montelukast. PGE2, paradoxically, is bronchodilatory when administered by inhalation, acting through EP2 receptors on airway smooth muscle — but this protective effect is lost in aspirin-exacerbated respiratory disease, where COX inhibition shunts arachidonic acid to the 5-LOX pathway, increasing leukotriene production.
Reproductive function: Prostaglandins play essential roles throughout reproduction. PGF2α produced by the uterine endometrium induces luteolysis — the regression of the corpus luteum — in the absence of pregnancy, re-initiating the estrous or menstrual cycle. During parturition, PGE2 and PGF2α promote cervical ripening and uterine contraction, and COX-2 expression in the fetal membranes increases dramatically in the hours preceding labor. PGE2 also maintains patency of the ductus arteriosus in the fetus — after birth, the abrupt increase in oxygen tension inhibits PGE2 production, allowing the ductus to close. In the male reproductive tract, PGE2 and PGF2α are present in seminal fluid and contribute to sperm motility and cervical mucus penetration.
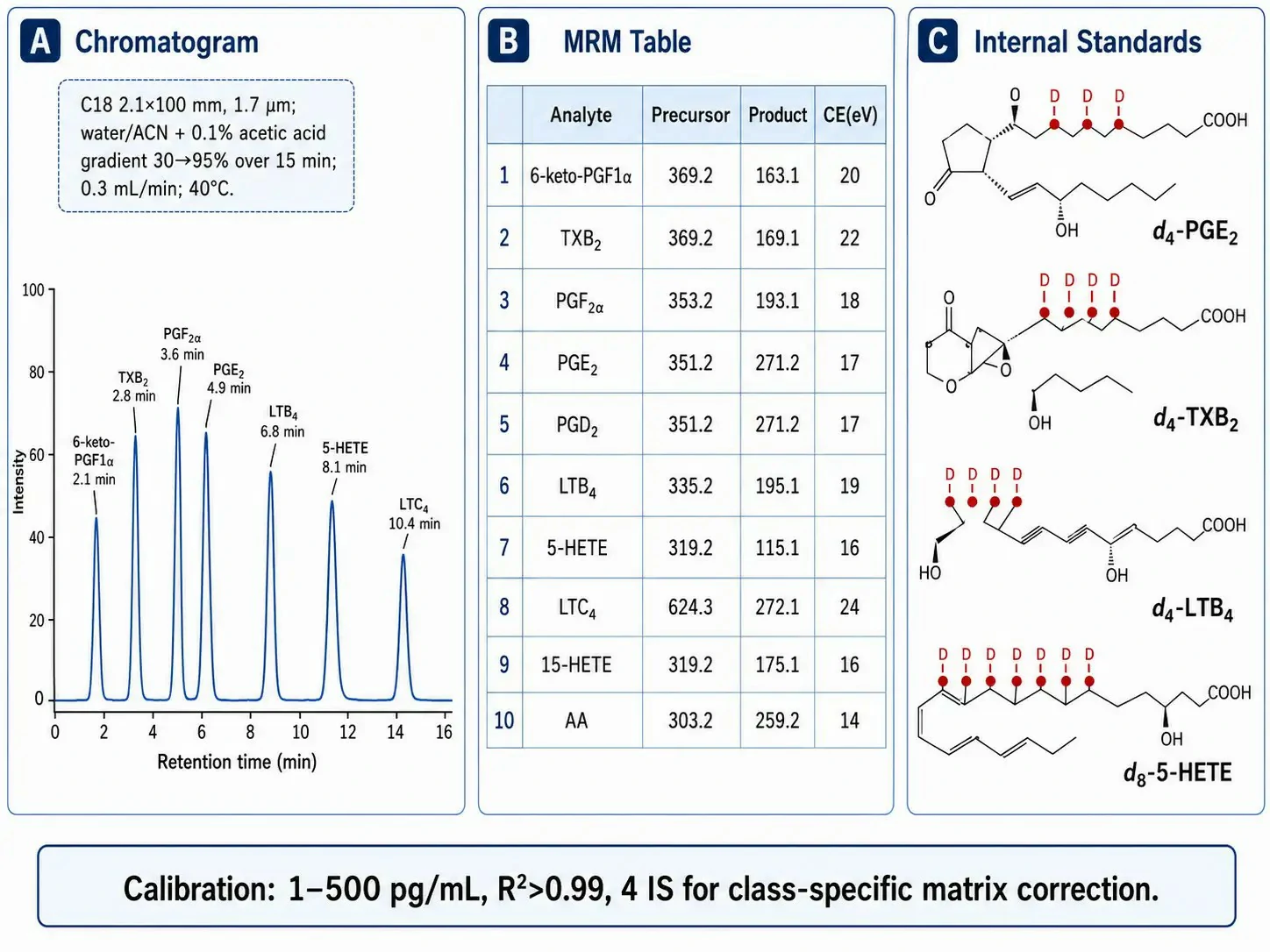
Figure 2: LC-MS/MS method parameters for eicosanoid profiling — MRM transitions, chromatography, and internal standard strategy
Eicosanoid Biosynthesis — Enzyme Selectivity and Analytical Implications
The three enzymatic pathways that convert arachidonic acid to eicosanoids differ in their product profiles, tissue distribution, regulation, and the analytical strategies required to measure their activity. Understanding these differences is essential for selecting the appropriate eicosanoid biomarkers and interpreting quantitative data from lipid mediator profiling experiments.
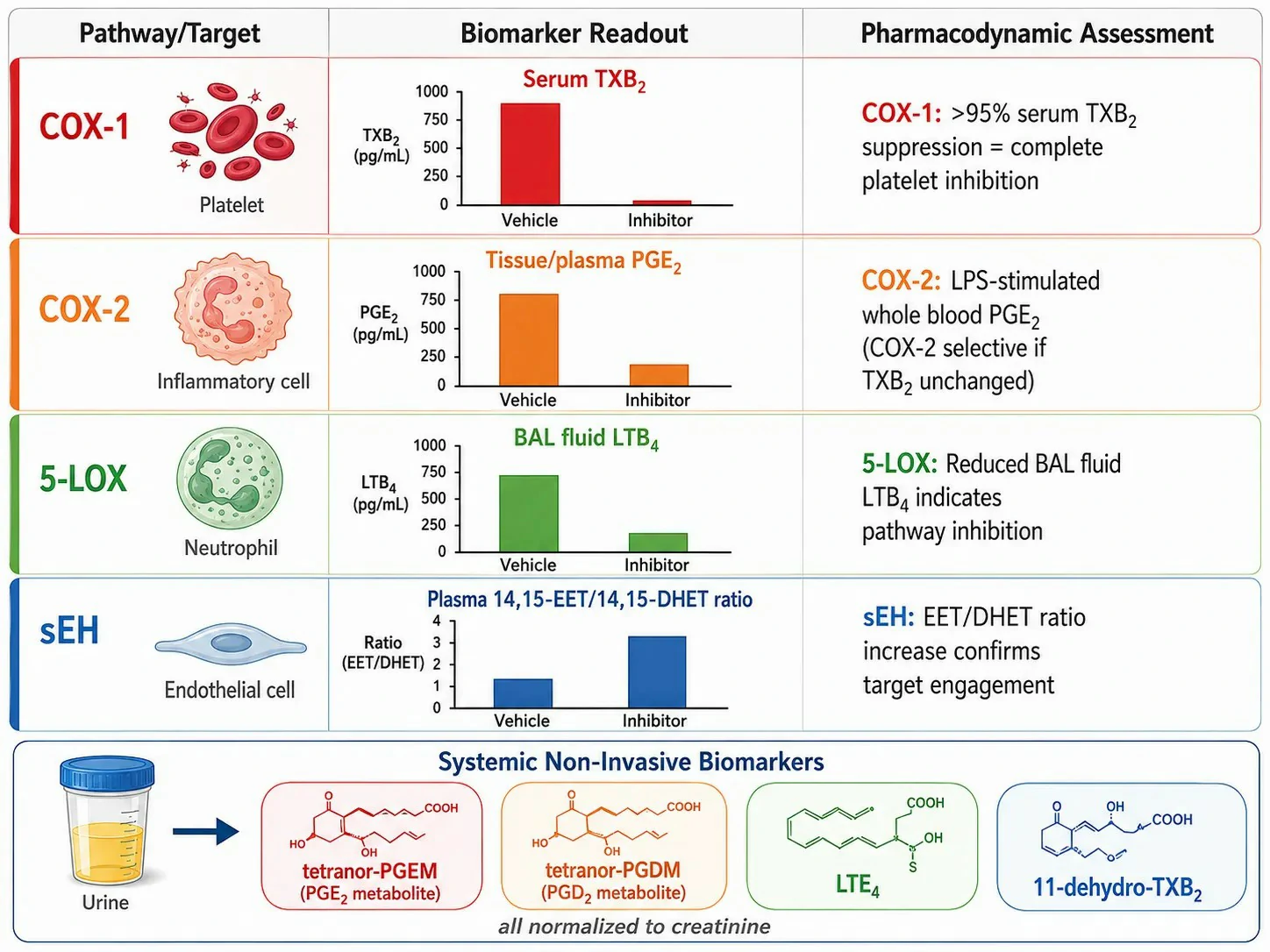
The cyclooxygenase pathway exists in two isoforms with distinct biological roles. COX-1 is constitutively expressed in most tissues and produces prostaglandins for physiological housekeeping — gastric cytoprotection (PGE2 and PGI2 protecting the gastric mucosa), platelet aggregation (TXA2 in platelets), and renal blood flow maintenance (PGE2 and PGI2 in the kidney). COX-2 is induced at sites of inflammation by cytokines (IL-1β, TNF-α), growth factors, and bacterial lipopolysaccharide. This differential expression provides a practical biomarker strategy: TXB2 in serum or as urinary 11-dehydro-TXB2 reports primarily on platelet COX-1 activity, while PGE2 in inflammatory exudates reports on COX-2 activity. In preclinical studies of COX-2 selective inhibitors, suppression of PGE2 at the inflammatory site with no change in serum TXB2 confirms COX-2 selectivity, while suppression of both indicates non-selective COX inhibition. COX-1 and COX-2 both convert arachidonic acid to PGG2 (via the cyclooxygenase active site) and then reduce PGG2 to PGH2 (via the peroxidase active site). The unstable PGH2 intermediate is subsequently converted to specific prostaglandins and thromboxane by cell-specific terminal synthases: TXA synthase in platelets (producing TXA2), PGI synthase in endothelial cells (producing PGI2), PGE synthase in inflammatory cells (producing PGE2), and PGD synthase in mast cells and the brain (producing PGD2).
The lipoxygenase pathway produces two functionally distinct families from a single unstable intermediate. 5-LOX, in concert with the 5-LOX activating protein (FLAP) which presents arachidonic acid to the enzyme, inserts molecular oxygen at the C5 position to form 5-HPETE, which is then dehydrated to the epoxide LTA4. LTA4 is the branch point: LTA4 hydrolase (a bifunctional zinc metalloprotease) converts LTA4 to LTB4 in neutrophils and macrophages, while LTC4 synthase conjugates LTA4 with glutathione to form LTC4 in mast cells, eosinophils, and basophils. LTC4 is exported from the cell by the multidrug resistance-associated protein MRP1 and is sequentially cleaved by extracellular γ-glutamyltransferase and dipeptidase to LTD4 and LTE4. The LTB4/LTC4 ratio in a biological sample reflects the relative contribution of the hydrolase and synthase pathways and identifies the predominant leukotriene-producing cell type — high LTC4/LTB4 ratios indicate mast cell and eosinophil activation, while high LTB4/LTC4 ratios indicate neutrophil-predominant inflammation. 12-LOX and 15-LOX produce 12-HETE and 15-HETE respectively, the latter serving as the precursor for lipoxin biosynthesis through transcellular pathways. Leukotriene profiling services quantify both LTB4 and cysteinyl leukotrienes to determine the cellular origin of LOX pathway activation.
The cytochrome P450 pathway produces two functionally opposed classes of eicosanoids. CYP epoxygenases (primarily CYP2C and CYP2J families) convert arachidonic acid to epoxyeicosatrienoic acids (EETs: 5,6-EET, 8,9-EET, 11,12-EET, and 14,15-EET), which are vasodilatory, anti-inflammatory, and cardioprotective mediators produced in the vascular endothelium. EETs are rapidly hydrolyzed to their corresponding dihydroxyeicosatrienoic acids (DHETs) by soluble epoxide hydrolase (sEH). The EET/DHET ratio is a pharmacodynamic biomarker for sEH inhibitor target engagement. CYP hydroxylases (primarily CYP4A and CYP4F families) produce 20-HETE, a potent vasoconstrictor that regulates renal and cerebral blood flow by inhibiting the large-conductance calcium-activated potassium channel in vascular smooth muscle. In preclinical models of hypertension, elevated 20-HETE production contributes to increased peripheral vascular resistance, and 20-HETE synthesis inhibitors reduce blood pressure.
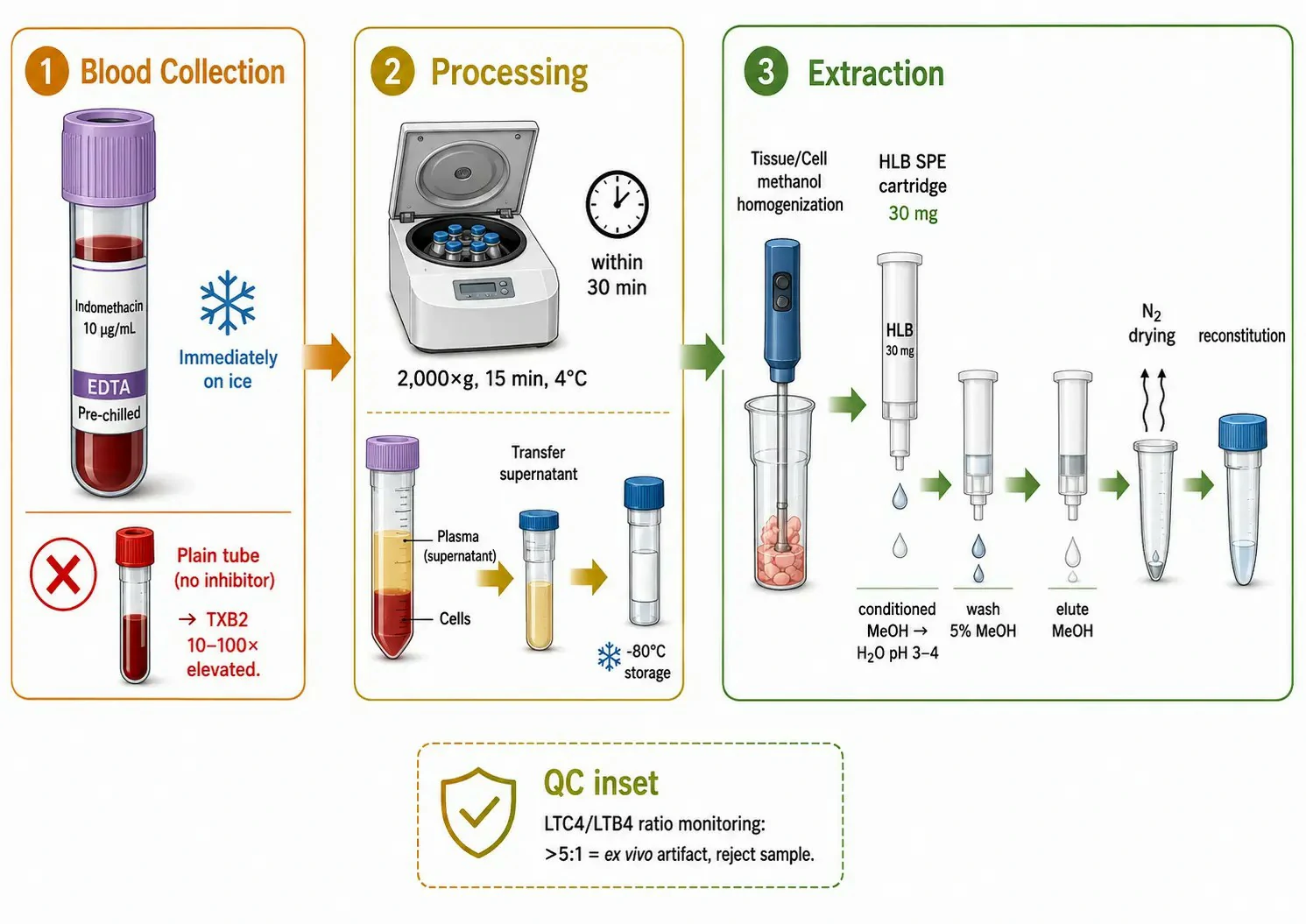
Figure 3: Sample collection and handling protocol for accurate eicosanoid quantification
LC-MS/MS Methods for Eicosanoid Profiling
Eicosanoid quantification by LC-MS/MS presents analytical challenges distinct from routine lipidomics. Circulating eicosanoid concentrations in biological fluids range from 5-50 pg/mL for TXB2 to 1-10 pg/mL for PGE2 and LTB4 — near the limit of quantification for many triple quadrupole instruments. Structural isomerism among prostaglandins (PGE2 and PGD2 share identical mass and MRM transitions and can only be distinguished chromatographically) requires high-resolution separations. Chemical instability (TXA2 t½ ~30 sec, measured as TXB2; PGI2 t½ ~3 min, measured as 6-keto-PGF1α) necessitates that the analytical target is the stable metabolite, not the biologically active parent compound.
Negative ion electrospray ionization (ESI⁻) is the standard detection mode for eicosanoids because the carboxylic acid group deprotonates efficiently at the acidic mobile phase pH (commonly 0.1% acetic acid). Each MRM transition must be individually optimized for collision energy. Representative transitions include: PGE2 and PGD2 (m/z 351→271, loss of H₂O + CO₂, CE 18-20 eV), TXB2 (m/z 369→169, CE 18 eV), 6-keto-PGF1α (m/z 369→163, CE 22 eV), LTB4 (m/z 335→195, CE 18 eV), LTC4 (m/z 624→272, CE 24 eV), 5-HETE (m/z 319→115, CE 16 eV), Resolvin D1 (m/z 375→141, CE 20 eV), 14,15-EET (m/z 319→219, CE 16 eV), and 14,15-DHET (m/z 337→207, CE 22 eV).
Chromatographic separation uses a C18 column (2.1×100 mm, 1.7 µm) with a water/acetonitrile gradient (both containing 0.1% acetic acid) at 0.3-0.4 mL/min and column temperature 40°C. The gradient starts at 30% acetonitrile and increases to 95% over 15 minutes, providing resolution of PGE2 (8.5 min) from PGD2 (9.2 min) with Rs ~1.3, sufficient for baseline separation. The column is washed with 95% acetonitrile for 5 minutes between injections to remove residual non-polar lipids. The minimum recommended set of deuterated internal standards is d4-PGE2, d4-TXB2, d4-LTB4, and d8-5-HETE, bracketing the major eicosanoid classes for class-specific matrix effect correction. Standards are added at the beginning of sample preparation and co-extract with endogenous eicosanoids through SPE. Calibration spans 1-500 pg/mL with R²>0.99 for all analytes. Eicosanoid LC-MS/MS quantification services provide deuterated IS for each major eicosanoid class with validated calibration ranges.
Sample collection and handling — controlling ex vivo artifacts: The most severe source of error in eicosanoid measurement is ex vivo formation during sample collection. Platelets express high levels of COX-1 and produce large quantities of TXA2 and 12-HETE within seconds of activation by contact with the collection tube, shear stress, or trace thrombin generated during clotting. A blood sample collected without precautions will have TXB2 concentrations 10-100-fold above the true circulating concentration.
The validated collection protocol requires blood drawn into pre-chilled EDTA tubes containing indomethacin (10 µg/mL, a COX inhibitor) or a commercial protease inhibitor cocktail with COX and lipase inhibitors, immediate placement on ice, centrifugation at 2,000×g for 15 min at 4°C within 30 minutes, and plasma storage at -80°C. Indomethacin in the collection tube reduces ex vivo TXB2 formation by >95% compared to EDTA alone. Serum, which is inherently an activated sample, reflects eicosanoid production capacity rather than circulating concentration and should be interpreted as a functional assay of platelet COX-1 activity. For tissue, the protocol requires liquid nitrogen freezing within 5 seconds of excision, pulverization in a liquid-N₂-cooled mortar, homogenization in ice-cold methanol containing 0.1% BHT (antioxidant) and 10 µM indomethacin, centrifugation, dilution to<10% 1="" 30="" and="" spe="" on="" polymeric="" reversed-phase="" cartridges="" conditioned="" with="" methanol="" equilibrated="" water="" at="" ph="" 3-4.="" eicosanoids="" are="" eluted="" ml="" dried="" under="" reconstituted="" in="" acetonitrile.="" the="" ltb4="" ratio="" serves="" as="" a="" quality="" control="" indicator="">5:1 in plasma suggests ex vivo leukotriene generation because LTA4 generated ex vivo by activated neutrophils can non-enzymatically conjugate with glutathione in red blood cells. Targeted metabolomics services provide validated protocols for eicosanoid extraction from plasma, serum, and tissue.
Eicosanoid Metabolism and Systemic Biomarkers
Eicosanoids are locally produced and locally acting mediators with short circulating half-lives. Their rapid metabolism serves a dual purpose: it prevents these potent signaling molecules from acting systemically when they are intended to regulate local processes, and it generates more stable metabolites that provide accessible windows into whole-body eicosanoid production.
The metabolic inactivation of eicosanoids proceeds through multiple routes. β-oxidation shortens the carboxyl-terminal carbon chain from the ω-end, producing dinor and tetranor metabolites that are two and four carbons shorter than the parent compound. ω-oxidation, catalyzed by CYP4 enzymes, introduces a hydroxyl group at the terminal methyl group, which is subsequently oxidized to a carboxyl group, creating a dicarboxylic acid that is more readily excreted. Conjugation reactions add glutathione (for leukotrienes, forming the cysteinyl leukotrienes and their further metabolites), glucuronic acid (by UDP-glucuronosyltransferases, producing more water-soluble glucuronides excreted in bile and urine), or taurine (for certain prostaglandins). These modifications collectively reduce biological activity and increase water solubility, facilitating renal and biliary excretion.
The metabolic pathway for PGE2 proceeds through sequential oxidation and reduction. 15-hydroxyprostaglandin dehydrogenase (15-PGDH) oxidizes the 15-hydroxyl group to a ketone, forming 15-keto-PGE2, which is the most rapid inactivation step. 15-keto-prostaglandin Δ13-reductase then reduces the 13,14-double bond to form 13,14-dihydro-15-keto-PGE2 (PGEM). PGEM undergoes β-oxidation from the carboxyl end and ω-oxidation from the methyl end to produce tetranor-PGEM, the major urinary metabolite of PGE2. PGD2 follows an analogous pathway to tetranor-PGDM, reflecting systemic PGD2 production primarily from mast cells. LTE4, the most stable cysteinyl leukotriene, is excreted in urine and provides a biomarker of systemic leukotriene production.
Urinary metabolites are present at 10-100-fold higher concentrations than their parent compounds in plasma and offer the analytical advantage of integrating eicosanoid production over the urine collection period rather than reflecting the instantaneous concentration at the time of blood sampling. Measurement requires β-glucuronidase treatment to hydrolyze glucuronide conjugates (20-50% of urinary metabolites), followed by SPE extraction and LC-MS/MS analysis using MRM transitions specific to the tetranor forms. Results are normalized to urinary creatinine. In preclinical studies of systemic inflammation, urinary tetranor-PGEM increases 5-20-fold within 6-12 hours of the stimulus and returns to baseline within 24-48 hours as inflammation resolves. Eicosanoid metabolic profiling services include urinary metabolite panels for systemic biomarker studies.
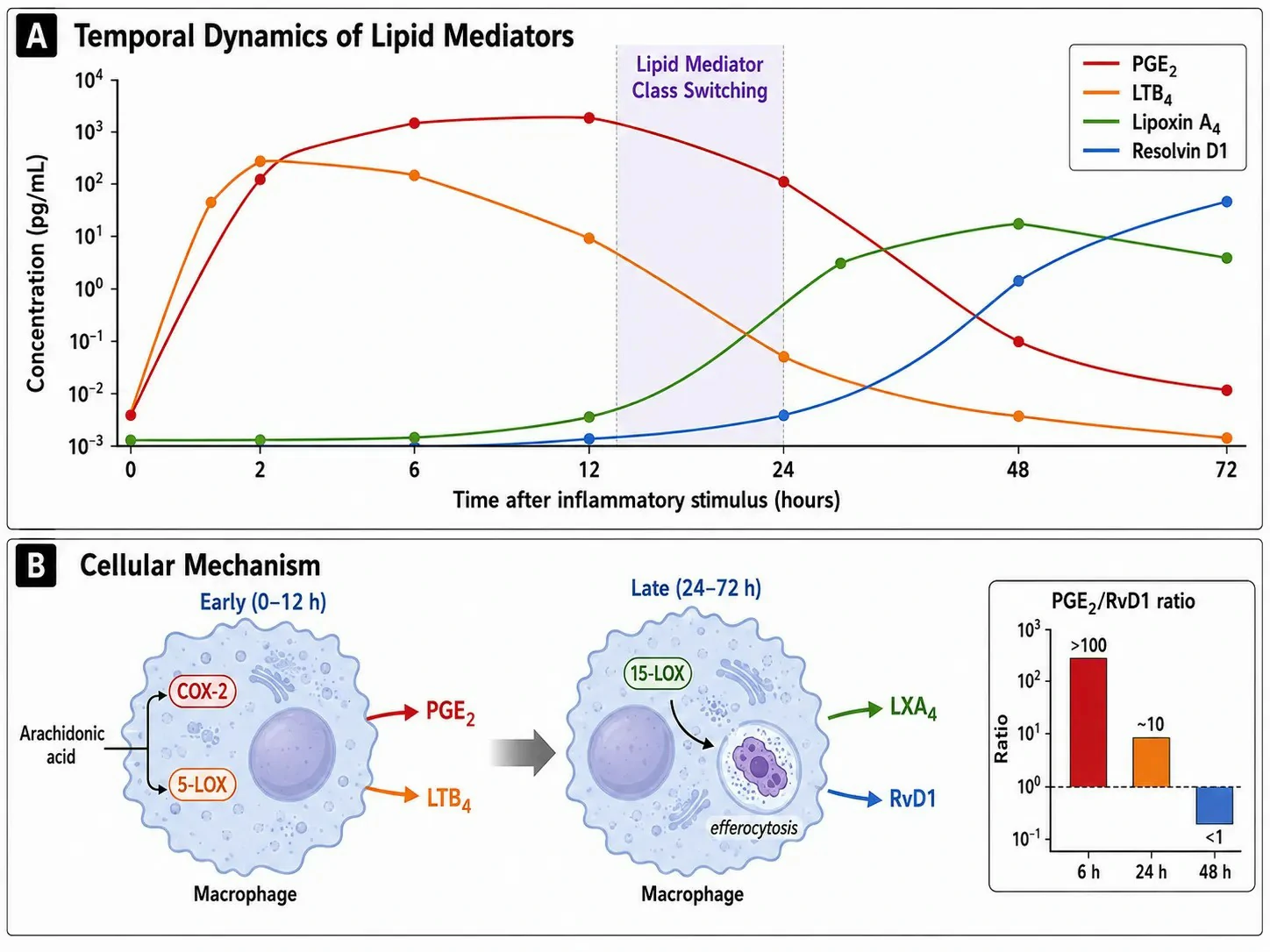
Figure 4: Lipid mediator class switching — from pro-inflammatory to pro-resolving mediators over the time course of inflammation
Lipid Mediator Class Switching — From Pro-Inflammatory to Pro-Resolving
Inflammation is not a static upregulation of pro-inflammatory mediators followed by passive decay. It is an active transition in which the initial phase — dominated by PGE2 and LTB4 — gives way to a resolution phase driven by a distinct set of SPMs. This transition, termed lipid mediator class switching, is essential for a return to tissue homeostasis and is actively regulated at the transcriptional and enzymatic levels.
The switching mechanism operates through a change in the enzymes expressed by the cells at the inflammatory site. During the early phase (0-12 hours), COX-2 and 5-LOX predominate, producing PGE2 and LTB4 from arachidonic acid. As the inflammatory infiltrate matures (12-24 hours), neutrophils undergo apoptosis and are cleared by macrophages through efferocytosis. This engulfment process signals the macrophage to upregulate 15-LOX while downregulating 5-LOX, shifting the arachidonic acid product profile from LTB4 (a neutrophil chemoattractant) to lipoxin A4 (a promoter of neutrophil apoptosis and macrophage efferocytosis). Simultaneously, the macrophage begins to utilize EPA and DHA as substrates for resolvin, protectin, and maresin biosynthesis. Resolvin D1, derived from DHA through sequential 15-LOX and 5-LOX activity, becomes detectable at 24-48 hours and reaches peak concentrations of 10-100 pg/mL — 100-1,000-fold lower than peak PGE2 concentrations, yet biologically effective because SPM receptors have sub-nanomolar affinity.
The analytical implication is that a single-time-point measurement of eicosanoids cannot characterize the inflammatory state. A time-course study with at least 3-5 time points is required to capture the trajectory. The PGE2/Resolvin D1 ratio declines from >100 at 6 hours to<1 at 48 hours in resolving inflammation — a parameter that distinguishes stalled inflammation (persistently high PGE2/SPM ratio) from resolving inflammation (declining ratio) in preclinical models of chronic inflammatory disease. Lipidomics profiling services measure the full spectrum of pro-inflammatory and pro-resolving eicosanoids from a single sample to capture this temporal transition.
Dysregulated Eicosanoid Signaling in Disease
The tight regulation of eicosanoid biosynthesis means that dysregulation — whether through increased production, decreased metabolism, or altered receptor expression — contributes to the pathogenesis of diverse diseases. Quantitative eicosanoid profiling can identify the specific pathways that are dysregulated and guide the selection of pharmacological interventions.
Inflammatory disorders: In rheumatoid arthritis, synovial fluid shows markedly elevated PGE2 (from COX-2 in synovial fibroblasts and infiltrating macrophages) and LTB4 (from 5-LOX in neutrophils). The LXA4/LTB4 ratio in synovial fluid correlates inversely with joint erosion scores, consistent with insufficient pro-resolving mediator production contributing to chronicity. In inflammatory bowel disease, colonic mucosal PGE2 is elevated 10-50-fold, produced by COX-2 induced in intestinal epithelial cells and lamina propria mononuclear cells. In asthma, bronchoalveolar lavage fluid contains elevated cysteinyl leukotrienes, and the LTE4 concentration in urine reflects the severity of airway inflammation. Aspirin-exacerbated respiratory disease is characterized by overproduction of cysteinyl leukotrienes at baseline and a further increase after COX-1 inhibition, due to the removal of PGE2-mediated suppression of 5-LOX activity in mast cells and eosinophils.
Cardiovascular disease: The balance between TXA2 (platelet-derived, pro-thrombotic) and PGI2 (endothelial-derived, anti-thrombotic) determines thrombotic risk at sites of atherosclerotic plaque. Plaque rupture exposes tissue factor and collagen, activating the coagulation cascade and platelets, generating TXA2 that amplifies thrombus formation. The urinary 11-dehydro-TXB2 concentration reflects in vivo platelet activation and is elevated in patients with acute coronary syndromes. Atherosclerotic arteries show reduced PGI2 production due to endothelial dysfunction, shifting the TXA2/PGI2 balance toward thrombosis. In hypertension, increased vascular 20-HETE production contributes to vasoconstriction and elevated peripheral resistance, and urinary 20-HETE is elevated in salt-sensitive hypertension models. EETs, produced by endothelial CYP epoxygenases, are vasodilatory and cardioprotective — they activate large-conductance calcium-activated potassium channels in vascular smooth muscle, causing hyperpolarization and relaxation. The EET/DHET ratio, reflecting the balance between EET production and sEH-mediated hydrolysis, is emerging as a biomarker of vascular health, with lower ratios indicating increased EET degradation and reduced vasodilatory capacity. Pharmacological sEH inhibition increases the EET/DHET ratio and reduces blood pressure in preclinical hypertension models, making this ratio a target engagement biomarker for sEH inhibitors in cardiovascular drug discovery. Eicosanoid profiling services include EET and DHET quantification for vascular function studies.
Cancer: COX-2 is overexpressed in many solid tumors, including colorectal, breast, lung, and prostate cancers. The resulting PGE2 production contributes to tumor progression through multiple mechanisms: it promotes tumor cell proliferation and survival, stimulates angiogenesis by inducing VEGF expression, and creates an immunosuppressive tumor microenvironment by inhibiting dendritic cell maturation, suppressing CD8+ T cell proliferation, and promoting regulatory T cell differentiation. Tumor tissue PGE2 concentration measured by LC-MS/MS correlates with COX-2 expression levels. In preclinical cancer models, COX-2 inhibition reduces tumor growth in COX-2-expressing tumors, and the pharmacodynamic readout is the reduction in tumor PGE2 concentration. Eicosanoid production by different cell types within the tumor microenvironment further complicates the profile — tumor-associated macrophages contribute PGE2 and LTB4, while the tumor cells themselves express varying profiles of COX-2 and downstream prostaglandin synthases depending on the tumor type. Prostaglandin analysis services provide quantitative PGE2 measurements for tumor tissue and the tumor microenvironment.
Figure 5: Eicosanoid biomarker strategies in disease research and preclinical drug discovery
Method Comparison — LC-MS/MS vs Immunoassay and GC-MS for Eicosanoid Measurement
The choice of analytical platform for eicosanoid measurement involves trade-offs among specificity, throughput, and the breadth of the eicosanoid panel. Each method occupies a specific niche in the preclinical research workflow.
Enzyme-linked immunosorbent assay (ELISA) is the most widely used method because of its low barrier to entry — it requires only a plate reader — and its throughput (40 samples in duplicate in 3-4 hours). Kits are commercially available for PGE2, TXB2, LTB4, 6-keto-PGF1α, and LTE4. However, ELISA suffers from cross-reactivity inherent to antibody-based detection: commercial PGE2 ELISA kits typically show 10-20% cross-reactivity with PGE1 and PGE3, which are present in plasma at concentrations that can reach 10-30% of PGE2 levels. The ELISA signal represents the sum of PGE2 plus a variable fraction of cross-reacting species, producing an overestimate that cannot be corrected without knowing the concentration of every interfering compound. For LTB4 ELISA, cross-reactivity with 6-trans-LTB4 and 12-epi-LTB4 — isomers produced by non-enzymatic hydrolysis of LTA4 — is a particular problem in samples where ex vivo leukotriene generation has occurred. Despite these limitations, ELISA remains useful for initial screening of COX pathway activation in large sample sets, with LC-MS/MS serving as the orthogonal confirmation method.
LC-MS/MS eliminates cross-reactivity by chromatographically separating isomeric eicosanoids before detection, and the MRM transition provides a second level of selectivity. A panel of 20-30 eicosanoids can be quantified in a single 15-minute run. The recommended workflow is ELISA screening → LC-MS/MS profiling of a representative subset of samples from each treatment group for confirmation and pathway-level interpretation.
Gas chromatography-mass spectrometry (GC-MS) was historically the primary method for eicosanoid analysis, particularly for urinary metabolites, and retains niche applications. GC-MS requires derivatization to make eicosanoids sufficiently volatile — methyl ester formation for the carboxyl group combined with trimethylsilyl ether or pentafluorobenzyl ester derivatization for hydroxyl groups. The derivatization adds 2-4 hours per batch and introduces variability from incomplete reaction. However, electron ionization spectra from GC-MS provide more structural information than CID spectra from triple quadrupole instruments and are useful for characterizing novel eicosanoid metabolites. GC-MS also offers chromatographic resolution superior to UPLC for some closely related isomers. LC-MS/MS has largely replaced GC-MS for routine eicosanoid quantification because it eliminates derivatization and provides comparable or better sensitivity on modern instruments, but GC-MS remains valuable for structural elucidation and for targeted analysis of urinary metabolites where the extensive sample preparation is already required for deconjugation and extraction. Targeted lipidomics services provide LC-MS/MS eicosanoid panels as the primary analytical platform and can incorporate GC-MS for novel metabolite characterization.
FAQ
What are the major classes of eicosanoids and which enzymes produce them?
Prostaglandins and thromboxanes (COX-1/COX-2), leukotrienes and lipoxins (5-LOX/12-LOX/15-LOX), EETs and HETEs (CYP epoxygenases and hydroxylases), and specialized pro-resolving mediators — resolvins, protectins, and maresins — derived from EPA and DHA rather than arachidonic acid.
How are eicosanoids measured by LC-MS/MS?
By negative ion ESI MRM on a C18 column with a water/acetonitrile gradient containing 0.1% acetic acid. Key transitions: PGE2 351→271, TXB2 369→169, LTB4 335→195, LTC4 624→272. Deuterated IS (d4-PGE2, d4-TXB2, d4-LTB4, d8-5-HETE) correct for extraction and matrix effects.
Why must blood samples be collected with special precautions for eicosanoid analysis?
Platelet activation during collection produces TXA2 and 12-HETE ex vivo, elevating concentrations 10-100-fold. Blood must be drawn into pre-chilled EDTA tubes containing indomethacin, kept on ice, and centrifuged within 30 minutes at 4°C.
What is lipid mediator class switching?
During acute inflammation, the initial phase produces pro-inflammatory PGE2 and LTB4 (0-12h). At 12-24h, macrophages clearing apoptotic neutrophils upregulate 15-LOX and utilize EPA/DHA to produce pro-resolving lipoxins, resolvins, and protectins (24-72h) that actively terminate inflammation.
How can eicosanoid metabolites in urine serve as systemic biomarkers?
PGE2 is metabolized to tetranor-PGEM and PGD2 to tetranor-PGDM, which are excreted in urine at 10-100-fold higher concentrations than parent compounds in plasma. After β-glucuronidase deconjugation, they are measured by LC-MS/MS and normalized to creatinine.
What is the difference between COX-1 and COX-2 inhibition and how is it measured?
COX-1 activity is measured as serum TXB2 (platelet COX-1). COX-2 activity is measured as LPS-stimulated whole blood PGE2 (monocyte COX-2). A compound reducing both is non-selective; a compound reducing PGE2 but not TXB2 is COX-2 selective.
References
- Dennis EA, Norris PC. Eicosanoid storm in infection and inflammation. Nature Reviews Immunology. 2015;15:511-523.
- Mosaad E, et al. The role(s) of eicosanoids and exosomes in human parturition. Frontiers in Physiology. 2020;11:594313.
- Johnson AM, Kleczko EK, Nemenoff RA. Eicosanoids in cancer: new roles in immunoregulation. Frontiers in Pharmacology. 2020;11:595498.
- Simultaneous profiling and quantification of 25 eicosanoids by LC-MS/MS. Metabolites. 2022;12:1203.
- Identification of specialized pro-resolving mediator clusters from human plasma. Scientific Reports. 2018;8:18050.











