Enzyme Activity and Reaction Mechanism Screening by MS — Label-Free Enzyme Kinetics, Inhibition Profiling, and Mechanism Elucidation
Measure the enzyme reaction directly: substrate mass, product mass, and reaction velocity — no fluorescent label, no coupled enzyme, no indirect proxy.
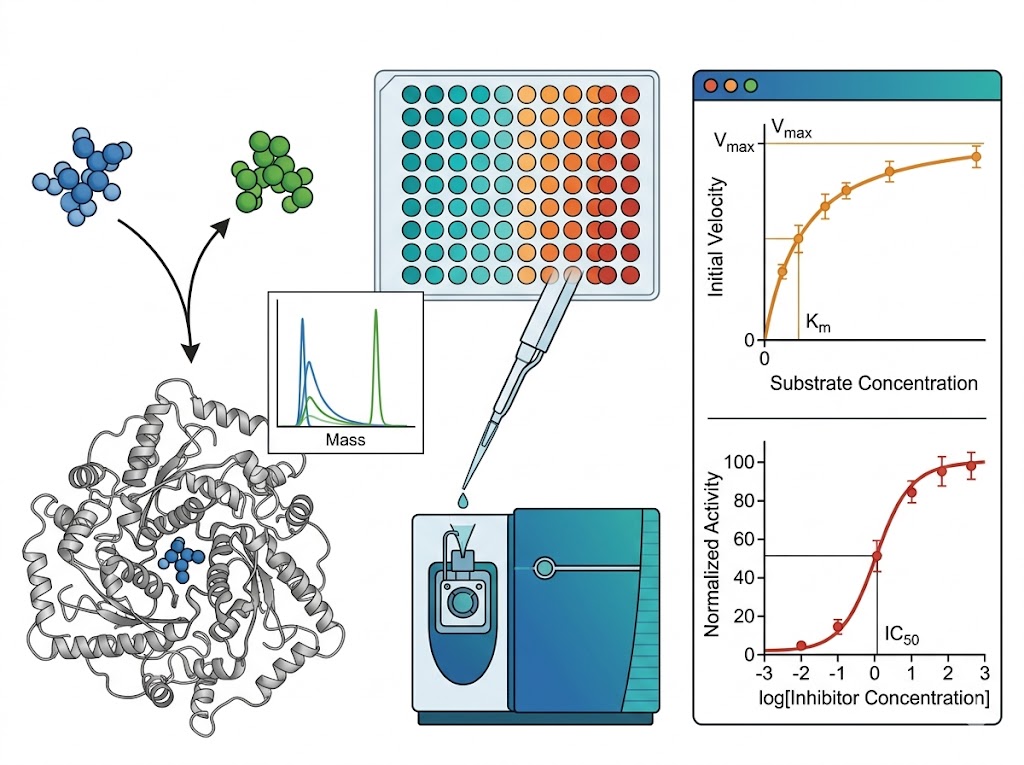
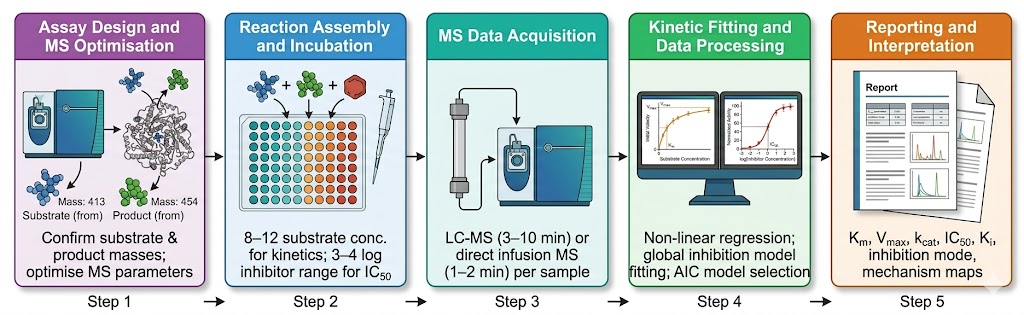
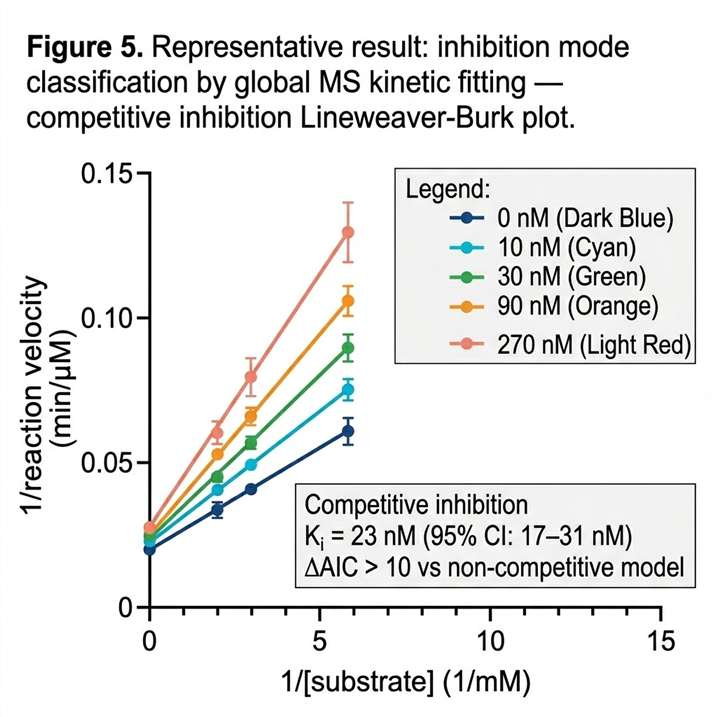
Enzyme activity measurement is fundamental to drug discovery — from primary inhibitor screening through lead optimisation to mechanism-of-action characterisation. Yet a substantial fraction of therapeutically relevant enzyme classes cannot be measured by fluorescence or absorbance-based assays. Methyltransferases, lyases, ligases, and isomerases lack chromogenic substrates. Coupled enzyme systems introduce artefacts, substrate limitations, and matrix incompatibility that compromise data quality. The limitation is not the enzyme — it is the detection method. Mass spectrometry-based enzyme activity screening circumvents this entirely: the mass spectrometer detects the substrate consumed and the product formed at their exact molecular masses, providing a direct, label-free measurement of enzyme activity that is independent of the optical properties of the molecules involved. Our enzyme activity and reaction mechanism screening service deploys high-resolution and triple-quadrupole MS platforms to deliver Michaelis-Menten kinetics, IC50 determination, inhibition mode classification, and reaction pathway characterisation — all from a single detection platform that reads the chemistry directly.
Key Advantages:
- Label-free detection of substrate and product by accurate mass — no fluorescent tag, chromogenic substrate, or coupled enzyme required.
- Applicable to every enzyme class: any enzyme whose substrate and product differ in mass is directly measurable.
- Crude lysate compatibility — unpurified enzyme preparations can be used for activity and inhibition measurements.
- Simultaneous kinetic and identity data — substrate consumption, product formation, and unexpected metabolites or intermediates are detected in the same experiment.
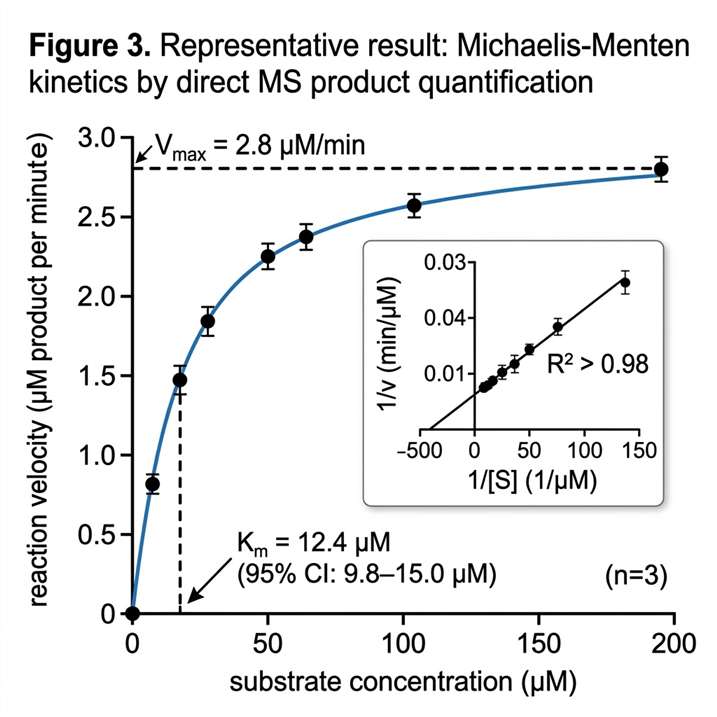
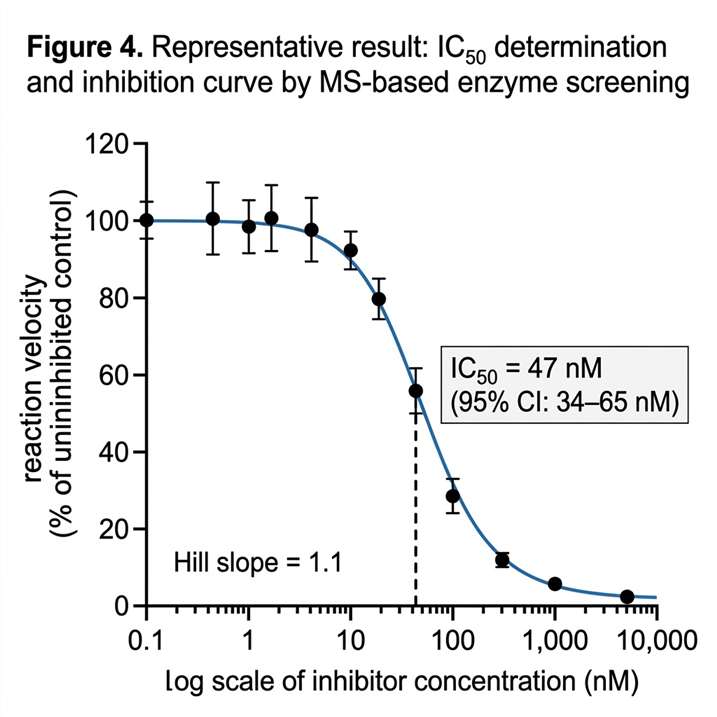
- Multi-parameter characterisation from a single platform — Km, kcat, IC50, inhibition mode, and reaction mechanism are accessible within the same MS workflow.
- Minimal assay development time — no probe engineering, no coupled enzyme optimisation, no fluorophore selection. If the substrate and product masses are known, the assay is defined.