Background
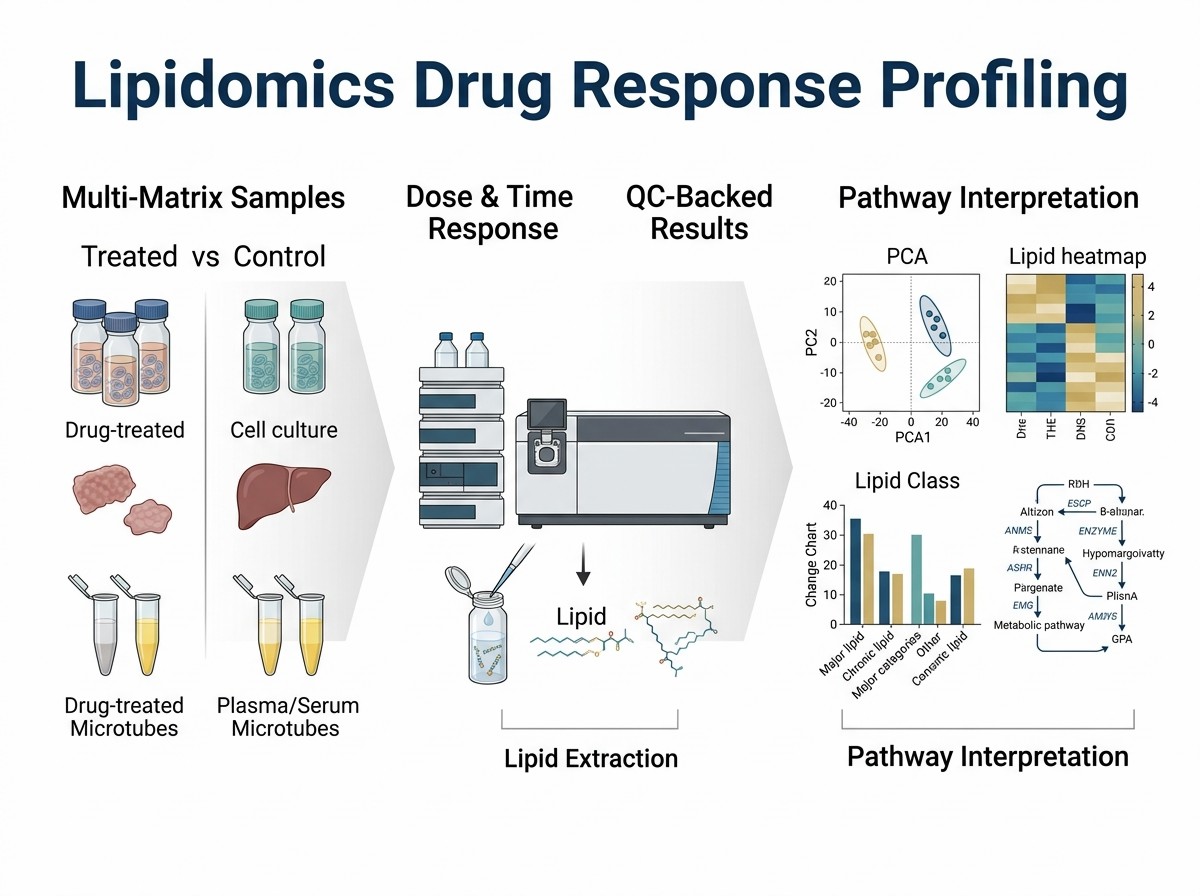
Drug resistance can involve changes in lipid metabolism, membrane biology, and lipid storage. In a 2025 study, Ramzy and colleagues investigated lipid signatures in FOLFOXIRI-naïve and FOLFOXIRI-resistant colorectal cancer cell models.
The study was designed to understand whether acquired resistance was associated with measurable lipidome changes. The authors used four human colorectal carcinoma cell lines and compared resistant clones with matched treatment-naïve cells.
Source: Identification of Lipid Species Signatures in FOLFOXIRI-Resistant Colorectal Cancer Cells
Methods
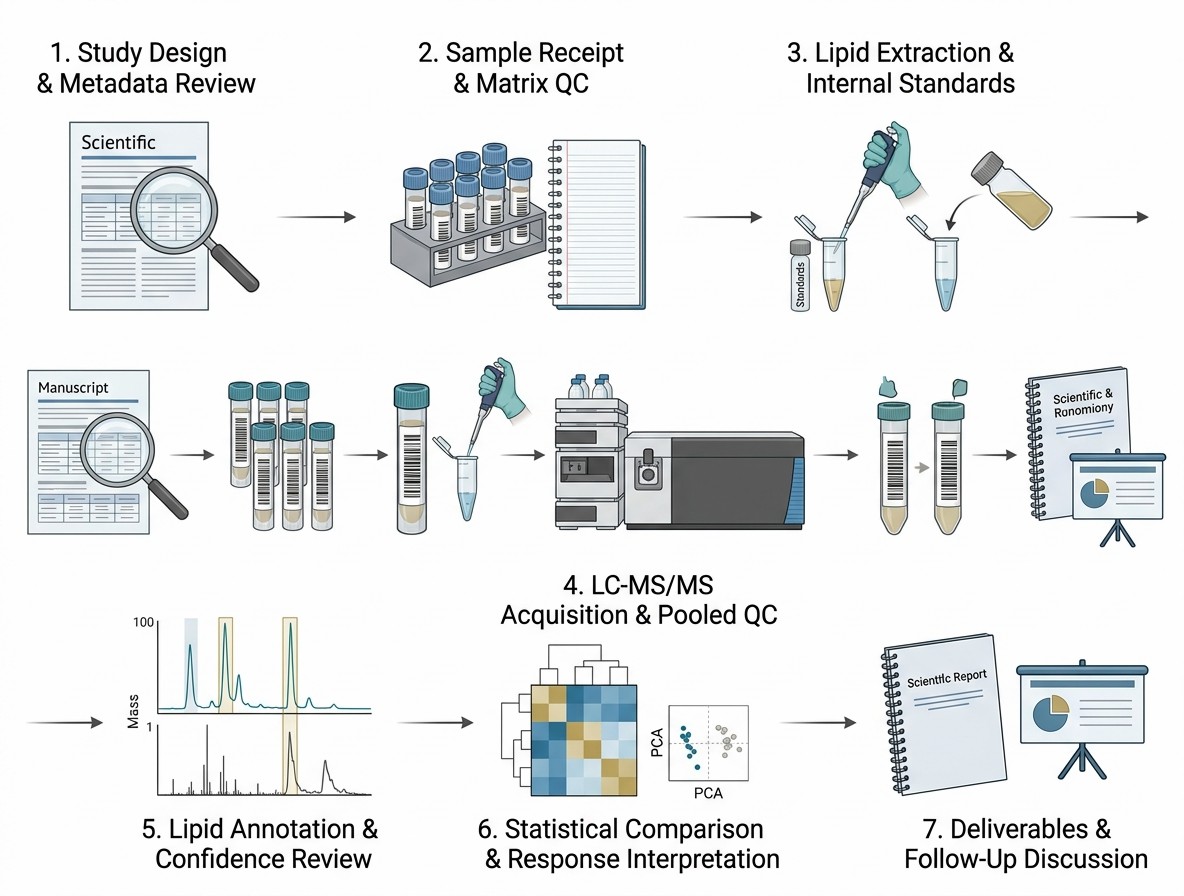
The researchers used untargeted LC-HRMS-based lipidomic profiling to compare lipid signatures across the cell models. They also used Analysis of variance-Multiblock Orthogonal Partial Least Squares, or AMOPLS, to separate variation linked to resistance, cell origin, and their interaction.
This design is relevant to drug-response lipidomics because it shows why treatment response should be interpreted in context. The lipidome shift was not only a simple resistant vs naïve pattern; it also depended on the cell line background.
Results
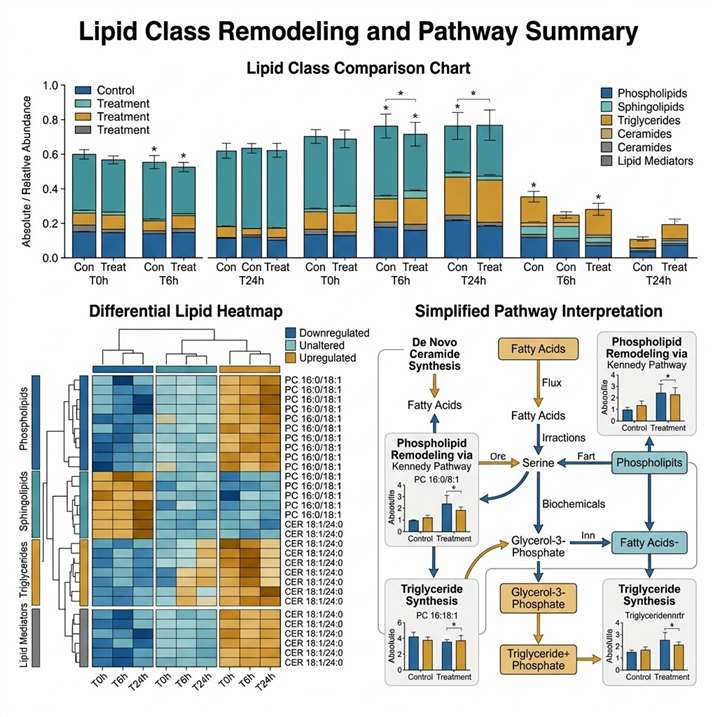
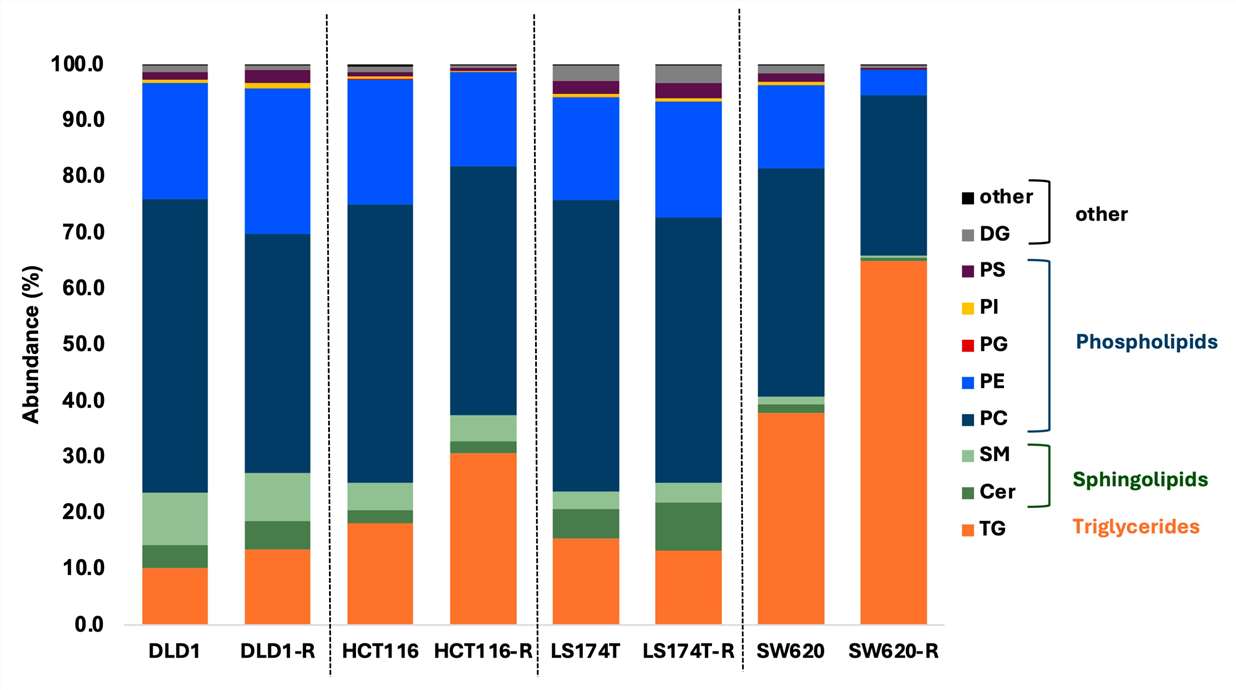
The paper reported 1737 MS2-matched lipids, which were further curated based on accurate mass, redundant adduct signals, and expected elution patterns. Figure 2 showed normalized distributions of the main lipid subclasses in FOLFOXIRI-naïve and FOLFOXIRI-resistant clones.
- HCT116 and LS174T cells showed similar lipid compositions, with 73% phospholipids, 7–8% total sphingolipids, and 15–18% triglycerides in the treatment-naïve state.
- DLD1 cells had the highest relative amount of sphingolipids among the four cell lines, at 14%.
- SW620 cells had the highest triglyceride proportion, at 38%, compared with 10–15% in the other three cell lines.
- DLD1-R, HCT116-R, and SW620-R resistant cells showed triglyceride abundance increases of 3%, 13%, and 27%, respectively.
- LS174T-R cells showed the opposite trend, with decreases in triglycerides and ether lipids.
- The authors reported that resistance-related lipid reprogramming was cell-line specific rather than a single universal lipid pattern.
Figure 2 from Ramzy et al. shows lipid subclass distribution shifts between FOLFOXIRI-naïve and FOLFOXIRI-resistant colorectal cancer cell models. The figure is a useful visual example of how lipidomics can reveal drug-response-associated lipid remodeling.
Conclusion
This study shows why lipidomics can be valuable for mechanism-focused drug response research. The lipid changes were not limited to one lipid class, and the response differed across cell backgrounds.
For drug discovery teams, this type of evidence can help connect treatment response with lipid pathway remodeling, identify model-specific differences, and select lipid signatures for follow-up testing.