Adenosine triphosphate (ATP) powers every step of genetic information transfer in the cell. From the unwinding of the DNA double helix by helicases to the peptide bond formation on ribosomes, from chromatin remodeling complexes that expose genes for transcription to the DNA repair machinery that preserves genomic integrity — each of these processes consumes ATP in specific, stoichiometric amounts that differ by orders of magnitude across pathways. Understanding these quantitative relationships is essential for interpreting cellular energy measurements, for distinguishing between adaptive metabolic changes and pathological ATP depletion, and for evaluating ATP-consuming enzymes as targets for preclinical drug discovery. The existing resource on this topic provides a comprehensive survey of the biological processes that depend on ATP. This guide extends that foundation by quantifying the ATP cost of each process, by connecting ATP consumption patterns to LC-MS/MS-based energy charge measurements, and by exploring how ATP-consuming enzymes serve as drug targets in preclinical research.
ATP quantitative analysis services provide validated LC-MS/MS methods for measuring ATP, ADP, and AMP with energy charge calculation in the context of DNA and protein synthesis studies.
Figure 1: The ATP cost of information transfer — quantitative comparison of ATP consumption per event across DNA replication, transcription, translation, and DNA repair
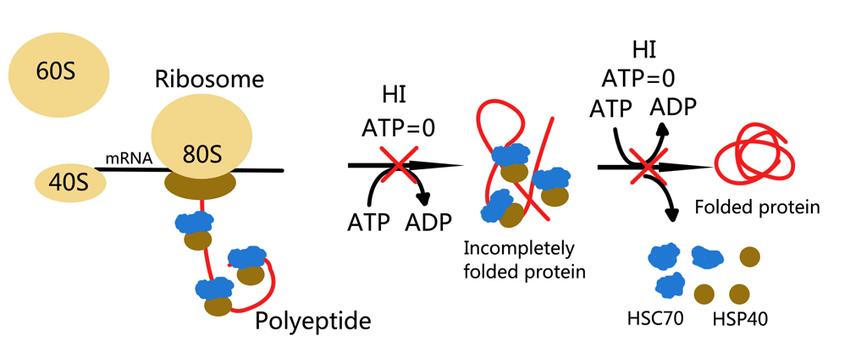
What the Existing Resource Covers — A Brief Recap
The existing resource on this topic provides detailed coverage of ATP's role across five biological domains. In DNA synthesis, it covers the ATP-dependent reactions catalyzed by DNA helicase (MCM complex separating the double helix), DNA polymerases α, δ, and ε (strand elongation), primase (RNA primer synthesis), and DNA ligase (Okazaki fragment joining). In protein synthesis, it covers tRNA aminoacylation by aminoacyl-tRNA synthetases (charging tRNAs with their cognate amino acids), translation initiation involving eIF4E and eIF4G, elongation driven by EF-Tu and EF-G with GTP hydrolysis, and termination by release factors. In cellular replication, it covers the coordination of S-phase DNA synthesis with protein synthesis, mitotic spindle assembly powered by microtubule motor proteins, and cytokinesis driven by the actin-myosin contractile ring. In genetic stability, it covers base excision repair (BER), nucleotide excision repair (NER), homologous recombination (HR), and chromatin remodeling by SWI/SNF and ISWI complexes. In genetic regulation, it covers transcription initiation by TFIID and RNA polymerase II, and epigenetic modifications including DNA methylation (DNMT enzymes), histone acetylation (HAT enzymes), and histone methylation (HMT enzymes). This guide does not repeat the descriptive biology of these processes. Instead, it quantifies the ATP cost of each one.
The ATP Cost of Information Transfer — A Quantitative Framework
Each step in the flow of genetic information — from DNA replication through transcription to protein synthesis — consumes a characteristic number of ATP molecules per reaction event. These stoichiometries are not arbitrary; they reflect the thermodynamic requirements of bond formation, the kinetic proofreading mechanisms that ensure accuracy, and the regulatory steps that couple energy status to biosynthetic rate.
For DNA replication, the incorporation of a single nucleotide into the growing strand costs approximately 2 ATP equivalents: one for the synthesis of the deoxynucleoside triphosphate (dNTP) from its diphosphate precursor by nucleoside diphosphate kinase, and one for the pyrophosphate released and hydrolyzed during the polymerase reaction itself. For a complete human genome of approximately 6×10⁹ base pairs (diploid), the total ATP cost of nucleotide incorporation alone exceeds 10¹⁰ ATP molecules. This does not include the ATP consumed by helicase activity (the MCM complex hydrolyzes 1-2 ATP per base pair unwound, adding another 6-12×10⁹ ATP), ligase activity (approximately 50 million Okazaki fragments requiring 1 ATP each for ligation), or the proofreading cycles of DNA polymerases δ and ε (each misincorporation excised by the 3'→5' exonuclease activity and re-synthesized costs an additional 1-2 ATP).
For protein synthesis, the ATP cost is even higher per information unit transferred. Charging a single tRNA with its cognate amino acid consumes 2 ATP equivalents (ATP → AMP + PPi, with pyrophosphate subsequently hydrolyzed to 2 Pi, making the net cost 2 high-energy bonds). Elongation factor EF-Tu delivers the charged tRNA to the ribosome and hydrolyzes 1 GTP per codon; elongation factor EF-G catalyzes ribosomal translocation and hydrolyzes another 1 GTP per codon. The total cost of incorporating one amino acid into a growing polypeptide chain is therefore approximately 4-5 ATP equivalents. For an average human protein of 400 amino acids, the synthesis cost is 1,600-2,000 ATP molecules — and this accounts only for translation, not for the subsequent ATP consumed by chaperone-assisted folding (Hsp70: 1 ATP per substrate binding-release cycle; Hsp90: 1 ATP per dimerization cycle; the TRiC/CCT chaperonin complex: approximately 130 ATP per substrate protein for the complete folding cycle within its central cavity).
For transcription, the cost is somewhat lower. Each ribonucleoside triphosphate (NTP) incorporated into RNA costs 1 ATP equivalent for its synthesis, and RNA polymerase II hydrolyzes the pyrophosphate released during each nucleotide addition, making the total cost approximately 1-2 ATP per nucleotide transcribed. A typical human gene of 10,000 base pairs therefore costs 10,000-20,000 ATP to transcribe once. However, regulatory processes associated with transcription — chromatin remodeling by SWI/SNF complexes (10-50 ATP per nucleosome repositioned), the assembly of the pre-initiation complex, and the phosphorylation of the RNA polymerase II C-terminal domain during promoter escape — add substantially to this baseline cost.
The relative magnitude of these costs has practical implications for cellular energy management. In rapidly proliferating cells such as activated lymphocytes and cancer cells, protein synthesis alone can account for 30-50% of total cellular ATP consumption, making it the single largest ATP sink after ion gradient maintenance. DNA replication, while individually costly per genome copy, is distributed across the approximately 8 hours of S-phase in human cells and represents a smaller fraction of the hourly ATP budget. However, the simultaneous demands of DNA replication and protein synthesis during S-phase create a peak ATP demand that must be met by coordinated upregulation of glycolysis and oxidative phosphorylation. When this coordination fails — for example, under hypoxia or mitochondrial dysfunction — the energy charge falls preferentially in S-phase cells, which are the most ATP-demanding subpopulation in any proliferating tissue. Targeted metabolomics profiling can measure the ATP/ADP/AMP ratios across cell cycle phases to map these energy demands.
Figure 2: The DNA replication fork — ATP consumption at each enzymatic step
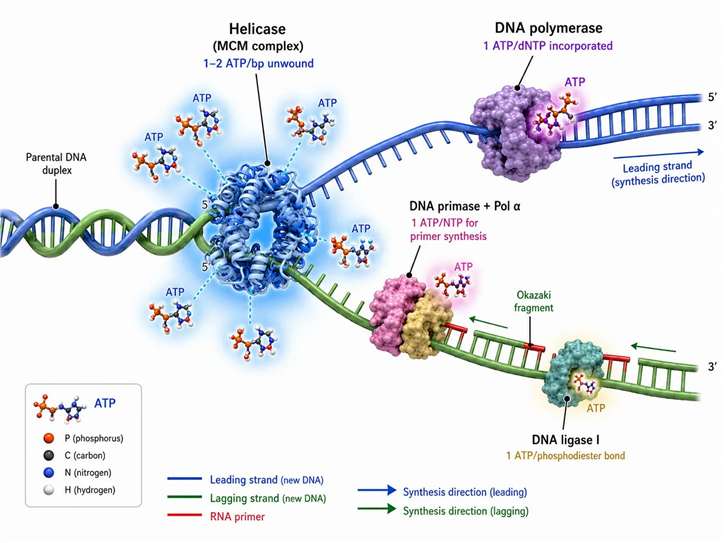
ATP as a Stoichiometric Fuel for DNA Replication
DNA replication is not a single ATP-consuming reaction but a coordinated sequence of ATP-dependent enzymatic steps, each with its own stoichiometry and regulatory logic. The MCM2-7 helicase complex, which unwinds the DNA double helix ahead of the replication fork, is the most ATP-intensive enzyme in the replisome. Each MCM hexamer hydrolyzes 1-2 ATP per base pair separated, and because the replication fork advances at approximately 50 nucleotides per second in human cells, a single active helicase consumes 50-100 ATP per second. With approximately 30,000-50,000 active replication origins firing during S-phase in a human cell, helicase ATP consumption at the peak of S-phase reaches approximately 1.5-5×10⁶ ATP per second across all forks combined.
Downstream of the helicase, DNA primase synthesizes short RNA primers (8-12 nucleotides) that provide the 3'-OH group required for DNA polymerase to begin strand elongation. Each primer nucleotide is an NTP that was synthesized from its diphosphate precursor at a cost of approximately 1 ATP equivalent per nucleotide. For the approximately 50 million Okazaki fragments synthesized on the lagging strand during human genome replication, primer synthesis alone consumes approximately 4-6×10⁸ ATP equivalents. DNA polymerase α extends these primers by 20-30 nucleotides before handing off to DNA polymerase δ (lagging strand) or ε (leading strand), each nucleotide addition costing 1 dNTP that represents approximately 1 ATP equivalent in synthesis cost.
The proofreading function of replicative DNA polymerases adds a variable but significant ATP cost. DNA polymerases δ and ε possess 3'→5' exonuclease activity that excises misincorporated nucleotides. The error rate without proofreading is approximately 10⁻⁴ to 10⁻⁵ per base pair; proofreading reduces this to approximately 10⁻⁶ to 10⁻⁷. Each proofreading cycle — excision of the mismatched nucleotide followed by re-insertion of the correct one — costs 1-2 additional ATP equivalents. For the entire human genome, with an intrinsic error rate of 10⁻⁵, approximately 60,000 proofreading events occur per replication cycle, consuming approximately 10⁵ ATP — a small fraction of the total replication ATP budget but essential for maintaining the mutation rate below the threshold that would compromise viability.
DNA ligase I, which seals the nicks between adjacent Okazaki fragments on the lagging strand, consumes 1 ATP per phosphodiester bond formed. The ATP is used to adenylate a lysine residue in the ligase active site, forming a covalent enzyme-AMP intermediate that activates the 5'-phosphate of the DNA nick for nucleophilic attack by the 3'-OH. With approximately 50 million Okazaki fragments per human genome replication, ligation consumes 5×10⁷ ATP — modest relative to helicase and polymerase costs but essential for completing replication. End-replication at chromosome termini, addressed by telomerase, adds a repeating TTAGGG sequence at a cost of 1 dNTP per nucleotide added, consuming ATP continuously at telomeres in cells that maintain telomerase activity.
Protein Synthesis and Quality Control — The Largest ATP Consumer
Protein synthesis is the single largest consumer of ATP among biosynthetic processes, and the reasons extend beyond the translation cycle itself. The 1,600-2,000 ATP molecules required to synthesize an average 400-amino-acid protein represent only the direct cost of tRNA charging and ribosomal elongation. The indirect costs — chaperone-mediated folding, quality control by the ubiquitin-proteasome system, and post-translational modifications — can add substantially to this baseline.
The molecular chaperone network is a major ATP consumer that is often overlooked in discussions of protein synthesis energetics. Hsp70, the most abundant and versatile chaperone in eukaryotic cells, hydrolyzes 1 ATP per substrate binding-and-release cycle. A protein that requires multiple Hsp70 binding cycles during its folding trajectory — not uncommon for large, multi-domain proteins — can consume 10-50 ATP in chaperone-assisted folding alone. Hsp90 operates on a different principle: ATP binding induces N-terminal dimerization, creating a molecular clamp that stabilizes client proteins in a folding-competent conformation, with 1 ATP consumed per clamp cycle. The TRiC/CCT chaperonin provides an enclosed folding chamber for approximately 10% of the eukaryotic proteome, consuming approximately 130 ATP per substrate protein across its ATP-driven conformational cycle. For a cell producing 10⁶ protein molecules per minute during peak growth, chaperone ATP consumption can rival the direct cost of translation.
Post-translational modifications (PTMs) add further ATP costs, though the mechanisms vary. Phosphorylation directly consumes 1 ATP per phosphate group transferred to a serine, threonine, or tyrosine residue by a kinase. A protein with 10 phosphorylation sites that is fully modified during its functional lifetime has consumed 10 ATP for modification alone — a small fraction of its synthesis cost but potentially critical for its regulatory function. Acetylation does not consume ATP directly; instead, acetyl-CoA — the acetyl donor for histone acetyltransferases (HATs) — is synthesized from acetate and CoA at a cost of approximately 1 ATP equivalent. For histones that are acetylated at 20-30 lysine residues across their N-terminal tails, this represents a significant ongoing ATP commitment. Methylation consumes S-adenosylmethionine (SAM) as the methyl donor; SAM synthesis from methionine and ATP costs approximately 12 ATP equivalents per molecule, making methylation one of the most expensive PTMs on a per-modification basis. PTM analysis services can map phosphorylation, acetylation, and methylation patterns to connect protein modification states with cellular ATP availability.
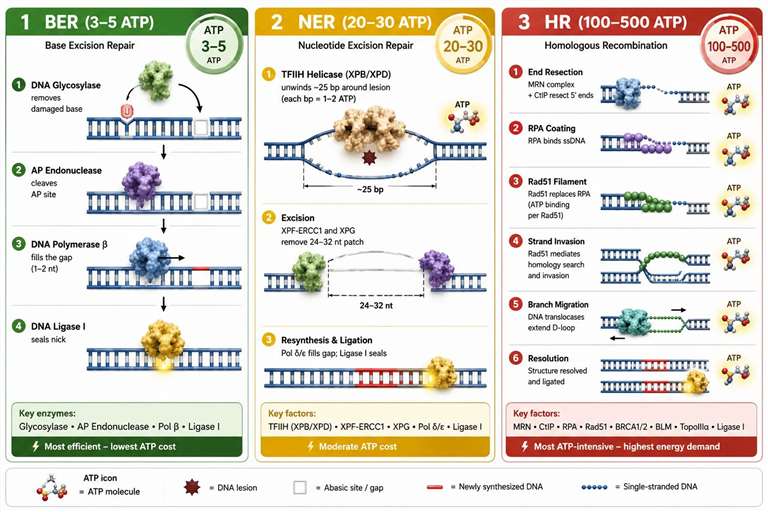
Figure 3: ATP cost comparison across DNA repair pathways — BER, NER, HR, and MMR
ATP-Driven Chromatin Remodeling and Epigenetic Energy Cost
The eukaryotic genome is packaged into chromatin, and access to DNA for transcription, replication, and repair requires ATP-dependent chromatin remodeling complexes to reposition or evict nucleosomes. The SWI/SNF family of remodelers, which includes the BAF and PBAF complexes in humans, uses the ATPase activity of the Brg1 or Brm catalytic subunit to translocate DNA around the histone octamer, consuming 10-50 ATP per nucleosome repositioning event. The ISWI and CHD family remodelers operate at a similar ATP cost but with different mechanistic outcomes — ISWI complexes primarily slide nucleosomes to create evenly spaced arrays, while CHD complexes are involved in nucleosome assembly and spacing during replication. The INO80 complex, which exchanges the H2A-H2B dimer for the H2A.Z variant, consumes ATP for both the remodeling and the histone exchange steps.
The epigenetic modifications that are deposited on histones and DNA also carry an ATP cost through their dependence on high-energy donors. DNA methylation by DNMT1 (maintenance methyltransferase) and DNMT3a/3b (de novo methyltransferases) uses SAM as the methyl donor. SAM synthesis from methionine and ATP proceeds through the methionine adenosyltransferase reaction, consuming the equivalent of approximately 12 ATP per SAM molecule produced. For a CpG island of 1,000 base pairs with 50% methylation density — 500 methylated cytosines — the initial methylation event has consumed the equivalent of 6,000 ATP just for SAM synthesis. Histone acetylation by HAT enzymes uses acetyl-CoA, whose synthesis from acetate, CoA, and ATP costs approximately 1 ATP equivalent per acetyl group. Histone methylation by HMT enzymes uses SAM at the same cost as DNA methylation. A nucleosome carrying histone H3 trimethylated at lysine 4, acetylated at lysine 9 and lysine 14, and trimethylated at lysine 27 has consumed ATP equivalents for each of these marks, representing an epigenetic energy investment that is maintained across cell divisions.
When ATP levels fall — during hypoxia, nutrient deprivation, or mitochondrial dysfunction — these ATP-dependent chromatin processes are among the first to be affected. Chromatin remodeling consumes ATP continuously for the dynamic regulation of gene expression; when ATP is limiting, remodeling slows, chromatin becomes less accessible, and the transcriptional response to environmental signals is blunted. The SAM/SAH ratio, which reflects the methylation potential of the cell, is coupled to ATP availability through the requirement for ATP in SAM synthesis. Measuring the SAM/SAH ratio alongside ATP and the energy charge by LC-MS/MS provides an integrated view of the cell's epigenetic capacity under different metabolic conditions.
ATP-Dependent DNA Repair — Energy Cost and Genomic Stability
DNA repair pathways differ dramatically in their ATP requirements, and this differential energy cost has consequences for repair pathway choice under conditions of energy stress. Base excision repair (BER), which corrects small base lesions such as oxidized or deaminated bases, is the least ATP-intensive pathway. The BER glycosylase (which recognizes and excises the damaged base), AP endonuclease (which cleaves the sugar-phosphate backbone at the abasic site), DNA polymerase β (which fills the single-nucleotide gap), and DNA ligase III (which seals the nick) together consume approximately 3-5 ATP per repair event. This low cost makes BER the default repair pathway for the thousands of spontaneous base lesions that occur in each cell per day, with each cell performing approximately 10,000-20,000 BER events daily at a total cost of 30,000-100,000 ATP per cell per day.
Nucleotide excision repair (NER), which removes bulky DNA adducts such as those caused by UV radiation and chemical carcinogens, is substantially more ATP-intensive at 20-30 ATP per repair event. The additional cost comes from the TFIIH helicase complex (containing the XPB and XPD ATPases), which unwinds approximately 25-30 base pairs of DNA around the lesion — each unwound base pair costing 1-2 ATP — and from the resynthesis of a 24-32 nucleotide patch by DNA polymerases δ or ε. The transcription-coupled NER sub-pathway, which preferentially repairs lesions on the transcribed strand of active genes, adds the ATP cost of the stalled RNA polymerase backtracking and the recruitment of the CSA and CSB proteins to the transcription bubble.
Homologous recombination (HR), the high-fidelity repair pathway for DNA double-strand breaks, is the most ATP-expensive repair mechanism at 100-500+ ATP per repair event. The cost derives from the extensive DNA processing required: the MRN complex (Mre11-Rad50-Nbs1) and CtIP resect the 5' ends of the break to generate 3' single-stranded DNA tails, consuming ATP for both the nuclease and helicase activities. RPA coats the ssDNA, Rad51 replaces RPA in an ATP-dependent filament assembly process, and the Rad51 nucleoprotein filament searches for and invades the homologous duplex — each Rad51 monomer hydrolyzing ATP during filament dynamics. The subsequent DNA synthesis, branch migration, and Holliday junction resolution each consume additional ATP. When the energy charge falls below approximately 0.70, HR is preferentially downregulated relative to non-homologous end joining (NHEJ), which is less ATP-intensive but more error-prone. This creates a direct mechanistic link between cellular energy status and mutagenesis: under energy stress, cells sacrifice repair fidelity for ATP conservation, increasing the mutation rate. ATP energy charge measurement in the context of DNA damage studies can reveal this repair pathway switching behavior.
ATP-Consuming Enzymes as Drug Targets in Preclinical Research
The ATP dependence of DNA and protein synthesis enzymes makes them vulnerable to pharmacological intervention at two distinct levels: direct inhibition of the enzyme's catalytic activity (competing with ATP at the active site or at allosteric regulatory sites) and indirect suppression through depletion of cellular ATP pools. Both strategies are actively pursued in preclinical drug discovery.
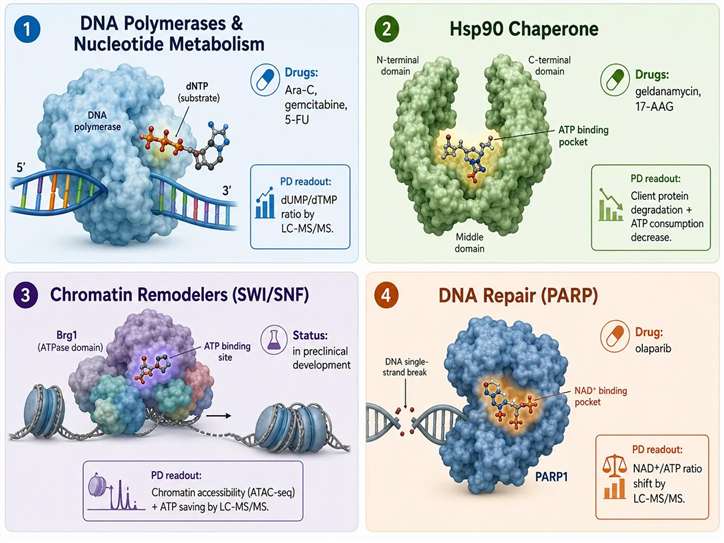
DNA polymerases and nucleotide metabolism enzymes represent the most established class of ATP-pathway drug targets. Nucleoside analogs such as cytarabine (Ara-C) and gemcitabine are phosphorylated to their active triphosphate forms by the same kinases that phosphorylate endogenous nucleosides, consuming cellular ATP in the process and producing modified nucleotides that inhibit DNA polymerases. Thymidylate synthase inhibitors such as 5-fluorouracil block the synthesis of dTMP from dUMP, causing dUMP accumulation, dTTP depletion, and replication fork stalling. The pharmacodynamic readout for these compounds — the dUMP/dTMP ratio measured by LC-MS/MS — directly reports on the blockade of an ATP-coupled nucleotide synthesis step. DHODH inhibitors (leflunomide, brequinar) block de novo pyrimidine synthesis at the dihydroorotate dehydrogenase step, which is directly coupled to the mitochondrial electron transport chain and therefore to ATP synthesis. The orotate/UMP ratio, measured by LC-MS/MS in cell or tissue extracts, serves as the PD biomarker for this class of compounds.
Hsp90 inhibitors represent a second class of ATP-pathway drug targets. Hsp90 is an ATP-dependent molecular chaperone that stabilizes a specific set of client proteins including steroid hormone receptors, kinases (HER2, BRAF, AKT), and transcription factors. Inhibitors such as geldanamycin and its derivatives (17-AAG, 17-DMAG) bind to the ATP-binding pocket in the N-terminal domain of Hsp90, preventing the ATP hydrolysis that drives the chaperone cycle. Without ATP, Hsp90 cannot complete its conformational cycle, and client proteins are degraded through the ubiquitin-proteasome pathway. Hsp90 is overexpressed 2-10-fold in many cancer cell types relative to normal tissue, and this overexpression makes cancer cells disproportionately dependent on Hsp90-mediated protein stabilization. The ATP consumption of Hsp90 in cancer cells — estimated at 10⁵-10⁶ ATP per cell per hour for the entire Hsp90 client protein network — represents a substantial fraction of the cancer cell's ATP budget that is not matched in normal cells, potentially contributing to the selectivity of Hsp90 inhibitors.
Chromatin remodeling complexes and epigenetic enzymes represent emerging ATP-pathway targets. The SWI/SNF ATPase subunits Brg1 and Brm are mutated in approximately 20% of human cancers, creating a dependence on the residual wild-type SWI/SNF activity that can be therapeutically exploited. Inhibitors of the Brg1 ATPase domain are in preclinical development, and their pharmacodynamic effect is measured by the decrease in chromatin accessibility at SWI/SNF-dependent enhancers — detected by ATAC-seq — combined with the decrease in total cellular ATP consumption attributable to SWI/SNF inhibition measured by LC-MS/MS. Similarly, inhibitors of DNA methyltransferases (DNMTi, such as azacitidine and decitabine) reduce the ATP cost of SAM synthesis for methylation reactions, and this metabolic sparing effect can be detected as a decrease in the cellular methionine consumption rate. Customized target engagement assays can measure both the direct ATPase inhibition and the downstream client protein depletion to fully characterize inhibitor pharmacology in the chromatin and epigenetic space.
Figure 4: ATP-consuming enzymes as drug targets in preclinical research
Analytical Perspective — Designing Experiments to Measure Synthesis-Linked ATP Consumption
Measuring the ATP cost of DNA and protein synthesis requires experimental designs that isolate the synthesis-related ATP consumption from the background of ongoing cellular metabolism. Three experimental strategies are most commonly employed.
First, the comparison of proliferating versus quiescent cells isolates the ATP consumption attributable to DNA replication and the associated increase in protein synthesis. Primary lymphocytes stimulated with mitogen (e.g., phytohemagglutinin) transition from G₀ to active proliferation over 24-48 hours, increasing their ATP consumption by 3-5-fold. LC-MS/MS measurement of ATP, ADP, and AMP at multiple time points during this transition, normalized to cell number, maps the temporal relationship between cell cycle entry and ATP demand. The energy charge should remain at 0.88-0.92 throughout the transition if the metabolic machinery adapts adequately; a decline in EC during the transition indicates that the proliferating cells are outrunning their ATP supply capacity.
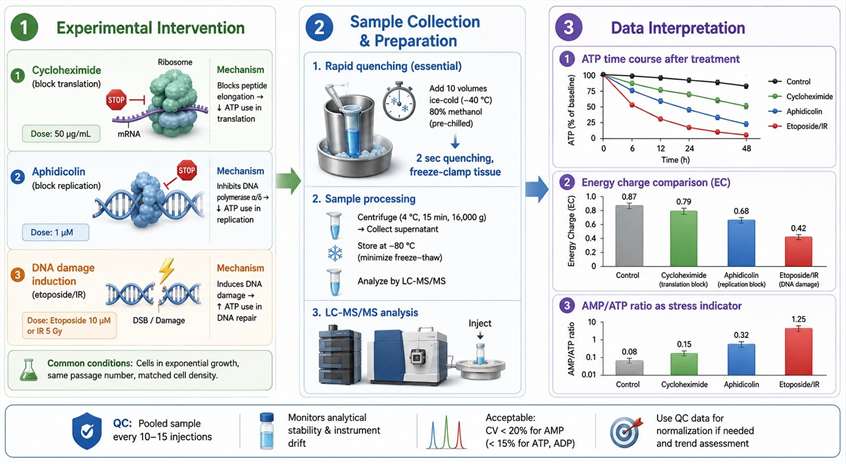
Second, the inhibition of specific synthesis processes with well-characterized inhibitors quantifies the ATP consumption attributable to that process. Cycloheximide blocks translation elongation by inhibiting the translocation step on the ribosome; the decrease in cellular ATP consumption after cycloheximide treatment — measured as the difference in ATP content between treated and untreated cells at 30-60 minutes after treatment — approximates the ATP cost of ongoing protein synthesis. Aphidicolin blocks DNA replication by inhibiting DNA polymerases α, δ, and ε; the ATP savings after aphidicolin treatment represent the replication-associated ATP consumption.
Third, DNA damage induction followed by LC-MS/MS time-course analysis measures the ATP cost of DNA repair. Treatment with etoposide (a topoisomerase II inhibitor that generates double-strand breaks by stabilizing the covalent topoisomerase-DNA intermediate) or with ionizing radiation produces a defined burden of DNA damage. The ATP consumption during the subsequent repair period — measured as a decrease in the energy charge or an increase in the AMP/ATP ratio relative to undamaged controls over 1-6 hours — reflects the repair ATP cost. This experimental design is particularly informative when comparing repair-proficient and repair-deficient cell lines, as the difference in ATP consumption between the two isolates the energy cost of the specific repair pathway that is functional in the proficient line. Metabolic flux analysis services can complement these ATP measurements by tracing the metabolic pathways that supply the additional ATP demand during replication and repair.
For all three experimental strategies, the sample preparation protocol must account for the rapid turnover of ATP described in detail in companion resources on ATP quantification. Briefly, cells must be quenched with cold 60% methanol (-40°C) within 2 seconds of harvest, tissues must be freeze-clamped within 5 seconds of excision, and the energy charge calculation should use isotopically labeled internal standards for ATP, ADP, and AMP individually to correct for differential extraction recovery. The interpretation of ATP consumption data in synthesis studies is most reliable when the energy charge is reported alongside the individual adenylate concentrations, as the EC provides a normalized readout that is less sensitive to variations in extraction efficiency than absolute ATP measurements. ATP energy charge measurement services provide the full adenylate panel with validated sample preparation protocols for DNA and protein synthesis studies.
Figure 5: Experimental design for measuring ATP consumption in DNA and protein synthesis studies
FAQ
How much ATP does it cost to replicate the entire human genome?
Approximately 10¹⁰ ATP molecules, including nucleotide synthesis, helicase unwinding, polymerase activity, and Okazaki fragment ligation. This cost is distributed across the approximately 8 hours of S-phase.
Which consumes more ATP in a cell — DNA replication or protein synthesis?
Protein synthesis — it accounts for 30-50% of total cellular ATP consumption in proliferating cells, compared to DNA replication which accounts for 5-10%. Translation is the single largest ATP sink after ion gradient maintenance.
Why does homologous recombination cost so much more ATP than base excision repair?
HR requires extensive DNA end resection by helicases and nucleases (50-100 ATP alone), Rad51 filament assembly with continuous ATP hydrolysis, strand invasion, branch migration of Holliday junctions, and gap-filling DNA synthesis (100-500 ATP total). BER repairs single-base lesions with only a glycosylase, AP endonuclease, short-patch polymerase, and ligase (3-5 ATP total).
How does ATP availability affect DNA repair pathway choice?
When ATP is limited (energy charge below 0.70), HR is preferentially downregulated, shifting double-strand break repair toward the less ATP-intensive but more error-prone NHEJ pathway. This directly links energy stress to increased mutation rates.
Can measuring ATP consumption identify the mechanism of a drug candidate?
Yes. A compound that reduces ATP levels specifically in S-phase cells but not in G₁ cells likely targets DNA replication. A compound that reduces ATP equally across cell cycle phases likely targets general metabolism or protein synthesis. LC-MS/MS ATP quantification across cell cycle-synchronized populations distinguishes these mechanisms.
How are Hsp90 inhibitors linked to ATP metabolism?
Hsp90 requires ATP to drive its chaperone conformational cycle. Hsp90 inhibitors (geldanamycin derivatives) bind to the ATP pocket of Hsp90, blocking ATP hydrolysis and causing client protein degradation. Cancer cells overexpress Hsp90 and are disproportionately dependent on its ATP-consuming activity.
References
- Mechanistic cross-talk between DNA/RNA polymerases and nucleotide metabolism. Nucleic Acids Research. 2020;48:8779-8796.
- ATP biosensor reveals microbial energetic dynamics across growth phases. Nature Communications. 2024;15:49579.
- How ATP and dATP act as molecular switches in ribonucleotide reductase. Biochemistry. 2024;63:2441-2452.
- Energy systems for ATP regeneration in cell-free protein synthesis. Methods in Molecular Biology. 2010;375:3-18.
- Measuring cellular adenine nucleotides by LC-MS. Methods in Molecular Biology. 2025;284:1-15.









