S-Nitrosation represents a nitric oxide (NO)-mediated post-translational modification (PTM) that modulates protein functionality through the conversion of cysteine (Cys) sulfhydryl (-SH) groups into S-nitrosothiol (-SNO) adducts. Antibodies, as pivotal agents in immunotherapy and diagnostic applications, are susceptible to functional alterations induced by S-nitrosation, including compromised structural integrity, impaired antigen-binding affinity, and diminished effector activities such as antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC). This review comprehensively examines the biological relevance of antibody S-nitrosation, evaluates current methodologies for its detection, and critically assesses mass spectrometry-based strategies for site-specific identification and functional characterization. Particular emphasis is placed on advancements in analytical techniques that elucidate the interplay between S-nitrosation dynamics and antibody performance, offering insights for both therapeutic optimization and mechanistic studies. For additional insights into detection protein nitrosation, explore our in-depth resource.
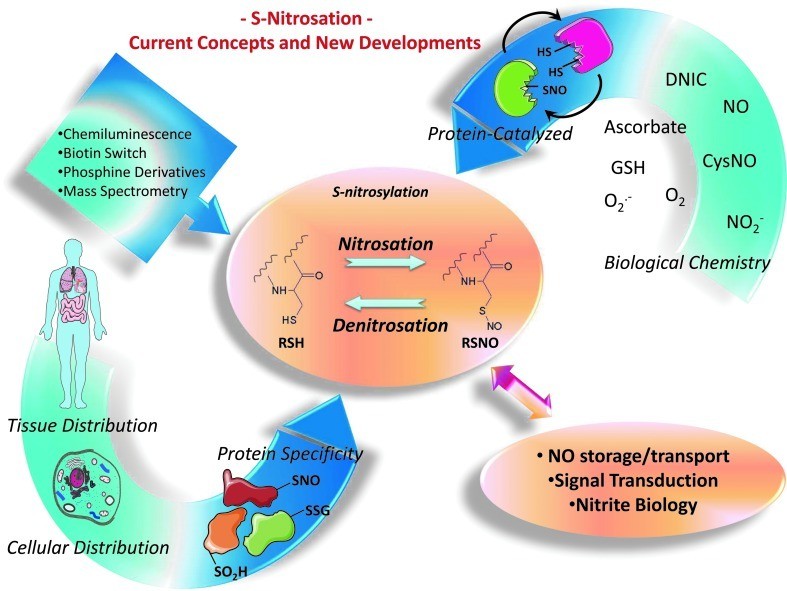 Several outstanding issues related to S-nitrosation (Thomas DD et al., 2012).
Several outstanding issues related to S-nitrosation (Thomas DD et al., 2012).
For additional insights into recombinant antibodies histone, explore our in-depth resource.
Systematic Mechanisms
S-Nitrosation, a dynamic post-translational modification (PTM), involves the covalent addition of nitric oxide (NO) to cysteine (Cys) residues via sulfhydryl group (-SH) conjugation, forming S-nitrosothiol (-SNO) adducts. This reversible modification serves as a critical regulator of antibody functionality, modulating immune responses through structural and functional adaptations. Below, we delineate its mechanistic framework across three dimensions: molecular foundations, regulatory networks, and functional outcomes. For additional insights into nitrosylation pathway, explore our in-depth resource.
1. Molecular Foundations of S-Nitrosation
Chemical Basis of Modification
- NO Biosynthesis: Generated by inducible nitric oxide synthase (iNOS) or endothelial nitric oxide synthase (eNOS), NO production surges in inflammatory or tumor microenvironments.
- Reaction Pathways:
- Direct oxidative coupling: Metal ions (e.g., Fe²⁺, Cu⁺) catalyze NO binding to Cys thiols, forming -SNO via radical intermediates.
- Indirect radical-mediated pathways: NO reacts with reactive oxygen species (e.g., O₂⁻) to yield peroxynitrite (ONOO⁻), which oxidizes thiols to -SNO.
Site-Specific Targeting Determinants
- Antibody Domain Preferences:
- Fc regions: Modification hotspots include CH2 domain Cys226 (IgG1) and hinge-region Cys220/Cys223 (IgG1/IgG4).
- Fab regions: Targets include complementarity-determining regions (CDRs) or framework residues (e.g., CDR3 in anti-PD-1 antibodies).
- Site Selectivity Factors:
- Local microenvironment: Low Cys pKa values and acidic residues enhance protonation, favoring modification.
- Structural exposure: Solvent-accessible Cys residues (e.g., hinge regions) are prioritized over buried disulfide-bonded sites.
2. Dynamic Regulation Networks
Modification Cycling: Writers and Erasers
- S-Nitrosation Induction:
- Enzymatic transfer: Thioredoxin reductase (TrxR) or protein disulfide isomerase (PDI) facilitates NO group transfer.
- Non-enzymatic diffusion: Elevated NO concentrations (e.g., in tumors) drive direct Cys modification.
- S-Nitrosation Removal:
- Reductive systems: Thioredoxin (Trx) and glutaredoxin (Grx) enzymatically reduce -SNO to restore -SH groups.
- Transnitrosylation: Glutathione (GSH) scavenges -SNO via thiol-disulfide exchange.
Redox Homeostasis and Environmental Influences
- ROS/NO Equilibrium: Reactive oxygen species (ROS) like H₂O₂ destabilize -SNO by oxidizing Cys thiols.
- pH Modulation: Acidic conditions (e.g., lysosomes, tumor stroma) promote Cys protonation, amplifying modification efficiency.
Crosstalk with Other Post-Translational Modifications
- Competitive Interactions: S-Nitrosation rivals S-glutathionylation and disulfide bond formation.
- Synergistic Relationships:
- Glycosylation: Fab-associated glycans may sterically shield adjacent Cys residues from modification.
- Phosphorylation: NO-cGMP-PKG signaling activates kinases, indirectly modulating antibody secretion and PTM dynamics.
3. Functional Consequences of S-Nitrosation
Impairment of Fc-Mediated Effector Functions
- FcγR Binding Disruption: Modification of CH2 domain Cys226 destabilizes Fc conformation, attenuating antibody-dependent cellular cytotoxicity (ADCC) and phagocytosis (ADCP).
- Complement Inhibition: -SNO adducts at C1q-binding sites (CH2 domain) reduce classical pathway activation.
Modulation of Antigen Recognition in Fab Regions
- Direct Antigen-Affinity Reduction:
- Conformational distortion: CDR3 modifications (e.g., in anti-PD-1 nivolumab) reduce structural rigidity.
- Charge interference: -SNO's negative charge disrupts antigen-antibody electrostatic interactions.
- Allosteric Effects: Fab conformational shifts indirectly alter adjacent antigen-binding pockets.
Impact on Antibody Stability
- Aggregation Propensity: -SNO-induced disulfide bond disruption promotes polymerization (e.g., IgG4 hinge-region modifications).
- Degradation Pathways:
- Ubiquitination-proteasomal degradation: Exposed Lys48-linked ubiquitination sites target modified antibodies for proteolysis.
- Lysosomal clearance: p62/SQSTM1 recognition directs S-nitrosated antibodies toward autophagic degradation.
Detection and Site-Specific Identification of S-Nitrosation
Biotin Switch Technique (BST)
- Principle: This method involves sequential chemical reactions to isolate S-nitrosated cysteine residues. Initially, unmodified sulfhydryl groups (-SH) are alkylated using methyl methanethiosulfonate (MMTS), while S-nitrosothiol (-SNO) moieties remain intact. Subsequently, ascorbic acid selectively reduces -SNO to regenerate -SH groups, which are then covalently tagged with biotin via a thiol-reactive reagent (e.g., HPDP-biotin). Biotinylated proteins are affinity-purified using streptavidin-coated beads, enabling detection via immunoblotting or site-specific characterization through mass spectrometry (MS).
- Experimental Workflow:
- Blocking free thiols: MMTS alkylates non-nitrosated cysteine residues.
- Selective reduction: Ascorbic acid converts -SNO groups to -SH.
- Biotin conjugation: HPDP-biotin labels newly exposed thiols.
- Enrichment and analysis: Streptavidin bead-based purification followed by Western blot or MS-based identification.
- Limitations:
- Low sensitivity for rare modifications may result in undetected sites, necessitating validation with high-resolution MS.
- Potential false positives due to incomplete blocking or nonspecific biotinylation.
Chemical Probes and Fluorescence-Based Detection
- CySNO Probes: Fluorescent chemical probes, such as Cy5-conjugated reagents, enable direct visualization of S-nitrosation dynamics. These probes chemically convert -SNO groups into stable fluorescent adducts, permitting real-time tracking of modification patterns and subcellular localization in antibodies using flow cytometry or confocal microscopy.
- Applications:
- Cellular imaging: Spatial resolution of S-nitrosated antibodies within live or fixed cells.
- Quantitative analysis: Fluorescence intensity correlates with modification levels, enabling semi-quantitative assessment.
- Advantages Over BST:
- Eliminates multi-step enrichment, reducing experimental complexity.
- Compatible with high-throughput screening and dynamic monitoring in intact cellular systems.
Services You May Be Interested In:
Additional Resources:
Mass Spectrometry
1. Sample Preparation and Modification Stabilization
Alkylation of Unmodified Thiols
- Reagent Optimization:
- MMTS: Selectively alkylates free cysteine thiols (-SH) without affecting S-nitrosothiol (-SNO) groups.
- NEM: Provides irreversible thiol blocking but requires compatibility with downstream reduction steps.
- Reaction Conditions: Performed in pH 7.4 buffer (e.g., 50 mM HEPES, 1 mM EDTA) at 25°C for 30 minutes, ensuring absence of reducing agents.
Stabilization and Labeling of S-Nitrosation
- Biotin Switch Technique (BST) Refinement:
- Selective Reduction: Low-dose ascorbic acid (1–5 mM) reduces -SNO to -SH while preserving disulfide bonds.
- Biotinylation: HPDP-biotin conjugates with regenerated thiols at pH 6.5–7.0, with labeling efficiency assessed via HPLC.
- Alternative Labeling Strategies:
- Click Chemistry: CuAAC links azide-functionalized -SNO to biotin probes, enhancing stability.
- Photoactivatable Probes: UV-crosslinking agents (e.g., Sulfo-SBED) capture transient modifications.
2. Antibody Digestion and Peptide Fractionation
Enzymatic Cleavage Strategies
- Multi-Enzyme Combinations:
- Trypsin + Glu-C: Enhances coverage of cysteine-dense regions (e.g., IgG1 hinge residues Cys220/Cys223).
- Lys-C + Chymotrypsin: Generates shorter peptides for hydrophobic CDR regions (e.g., anti-PD-1 antibodies).
- Accelerated Digestion:
- Microwave-assisted proteolysis reduces processing time (5 minutes vs. 16 hours) while preserving labile modifications.
- Ultrasonication disrupts antibody aggregates, improving enzymatic accessibility.
Peptide Enrichment Techniques
- Chromatographic Methods:
- SCX: Isolates positively charged SNO-modified peptides.
- TiO₂ Affinity: Captures acidic peptides via phosphate coordination.
- Microfluidic Systems: Nano-LC minimizes sample loss, boosting sensitivity for low-abundance modifications.
3. Mass Spectrometry Data Acquisition
Instrumentation and Parameterization
- High-Resolution MS Platforms:
- Orbitrap Exploris 480: Operated at 480,000 resolution (@200 m/z) to resolve isotopic fine structures.
- DDA Mode: Prioritizes cysteine-containing peptides via mass shifts (e.g., +78.98 Da for -SNO).
- Ion Mobility Enhancements:
- FAIMS: Separates isobaric peptides (e.g., SNO vs. phosphorylated forms).
- TIMS: Utilizes CCS values for structural discrimination.
Fragmentation Optimization
- ETD: Preserves labile -SNO groups during cleavage (10–20 ms reaction time, charge states z=2–3).
- Hybrid Modes: EThcD combines ETD and HCD to balance sequence coverage and modification localization.
4. Data Interpretation and Validation
Computational Analysis
- Database Search Parameters:
- Include dynamic modifications (+78.98 Da for -SNO, +15.99 Da for oxidation).
- Algorithm-Assisted Prediction: Tools like DeepSNO (CNN-based) predict modification-prone cysteines (NAR, 2021).
Experimental Verification
- Synthetic Peptide Standards: Validate retention times and fragmentation patterns (e.g., IgG1 Cys226 peptide).
- Quantitative Profiling:
- SILAC: Compares SNO levels in iNOS-overexpressing vs. control cells.
- TMTpro 16-plex: Multiplexed quantification across NO donor gradients.
5. Technical Challenges and Mitigation
Enhancing Low-Abundance Detection
- Immunoaffinity Enrichment: Anti-SNO antibodies (e.g., EMD Millipore) or engineered nanobodies (VHH) improve specificity.
- Signal Amplification: Proximity ligation assays (PLA) with DNA barcodes enhance detection limits.
Addressing Modification Instability
- In Situ Stabilization:
- Cryogenic MALDI-TOF reduces thermal decomposition.
- Rapid UV-crosslinking preserves transient modifications.
- Chemical Derivatization: Converts -SNO to stable sulfonic acid (-SOH) for robust analysis.
Site Assignment Accuracy
- CCS Libraries: Match experimental ion mobility values to theoretical predictions (e.g., Skyline software).
- Multistage MS: Orbitrap Astral enables MSⁿ fragmentation for resolving complex modification landscapes.
Functional Implications
1. Modulation of Immune Effector Functions
- ADCC Potentiation: Structural shifts in antibody Fc domains induced by S-nitrosation may augment binding capacity to FcγRIIIa (CD16) on natural killer (NK) cells. For instance, macrophage Septin2 nitrosylation modulates inflammatory responses via RAC1-NF-κB signaling, hinting at analogous mechanisms for optimizing antibody-effector cell interactions (Zhang Y ET AL., 2024).
- Complement Pathway Regulation: Modification of C1q-binding regions may alter classical complement activation. While S-nitrosation of UBE2D1 impedes ubiquitin ligase activity, analogous targeting of complement regulatory proteins could indirectly influence complement-dependent cytotoxicity (CDC) (Fujikawa K et al., 2020).
2. Bidirectional Antigen-Binding Modulation
- Structural Plasticity Alters Binding Strength: Disulfide rearrangement or electrostatic changes in Fab regions due to S-nitrosation may enhance or impair antigen engagement. For example, HCMV pp65 nitrosylation targeting C276/C306 residues diminishes cGAS antagonism, a principle potentially applicable to antibody variable regions (Cox JB et al., 2025).
- Epitope Occlusion: Steric hindrance adjacent to antigen-binding domains (e.g., CDR regions) may directly block recognition, reducing neutralization efficacy (Cox JB et al., 2025).
3. Stability and Metabolic Regulation
- Extended Circulatory Persistence: By attenuating proteasomal degradation, S-nitrosation may prolong antibody half-life. GSNOR-mediated S-nitrosothiol equilibrium attenuates ubiquitin-dependent proteolysis, suggesting analogous protection against intracellular protease recognition.
- Oxidative Resilience: In inflammatory settings, antibody nitrosylation may neutralize reactive oxygen species (ROS), preserving functionality. For example, DHAR nitrosylation in plant systems enhances ascorbate recycling, a mechanism potentially conserved in antibodies to resist oxidative inactivation.
4. Microenvironment-Driven Adaptation
- Vascular Dynamics and Tissue Penetration: S-Nitrosation regulates vascular endothelial function (e.g., via NO signaling), influencing antibody enrichment at disease sites. H2S-mediated S-nitrosation suppresses vascular smooth muscle proliferation, a mechanism that may indirectly enhance antibody infiltration into inflamed tissues.
- Immune Evasion Modulation: While viral protein nitrosylation (e.g., HCMV pp65) weakens immunosuppression, host antibody modifications might enhance pathogen surveillance or inhibit immune checkpoints (e.g., PD-1/PD-L1), balancing immune escape (Cox JB et al., 2025).
5. Therapeutic Applications
- Precision S-Nitrosation Engineering: Pharmacological GSNOR suppression (e.g., N6022) amplifies ADCC efficacy, while RAC1 inhibitors (e.g., NSC23766) mitigate hyperinflammatory risks, optimizing therapeutic windows.
- Resistance Reversal: In HER2-targeted therapies, GSNOR deficiency-linked SNO imbalance correlates with resistance. Restoring SNO equilibrium emerges as a resistance-countering strategy, potentially resensitizing tumors to treatment (Bignon E et al., 2018).
Applications
- RELAX10 enhances NO signal to mediate: RELAX10 activates AKT through RXFP1 receptor, promotes eNOS phosphorylation (Ser1177) and increases NO synthesis. ISO activates Ca²⁺- calmodulin-dependent eNOS activity through β -adrenergic receptor, and further releases NO. NO modifies the target protein through nitrosation (-SNO) of sulfhydryl (-SH) and regulates its function. This process depends on NO concentration and redox state (such as GSH/GSSG ratio) in microenvironment. The amplification of NO signaling pathway may protect myocardial function by modifying more target proteins (Sun J et al., 2019).
- The mechanism of drug resistance of trastuzumab driven by SNO homeostasis imbalance in HER2 positive breast cancer: In HER2-positive breast cancer, trastuzumab resistance is strongly associated with disrupted S-nitrosation (SNO) homeostasis. Inhibition of S-nitrosoglutathione reductase (GSNOR) diminishes trastuzumab's anti-proliferative and pro-apoptotic efficacy, primarily through activation of pro-survival signaling cascades such as AKT, Src, and c-Abl. Additionally, S-nitrosation of apoptosis regulators like Apaf-1 and c-Jun further suppresses cell death pathways. Clinical evidence reveals that reduced GSNOR expression correlates with tumor advancement in HER2-positive patients, whereas elevated ADH5/GSNOR gene levels predict improved survival outcomes. These findings highlight the potential of restoring SNO balance or pharmacologically targeting GSNOR to counteract resistance mechanisms, offering novel strategies to enhance anti-HER2 therapy by modulating critical survival and apoptotic pathways (Cañas A et al., 2016).
- TrxR regulates the balance of SNO in ER+breast cancer and promotes the phenotype and proliferation of tumor stem cells: In estrogen receptor-positive (ER+) breast cancer, thioredoxin reductase (TrxR) critically modulates S-nitrosation (SNO) equilibrium. Disrupting SNO homeostasis—via TrxR inhibition or exogenous SNO supplementation—selectively enhances ER+ MCF-7 cell proliferation and survival, a process governed by ERα signaling and p53 functionality. Mechanistically, this involves Akt/Erk1/2 pathway activation, cyclin D1 upregulation, and induction of cancer stem cell phenotypes, including CD44+ subpopulation expansion and p53-dependent reprogramming. Clinically, reduced TrxR1 (TXNRD1) expression in ER+ tumors, particularly those retaining wild-type p53, correlates with disease progression. These findings implicate SNO imbalance as a driver of ERα/TrxR1/p53 axis dysregulation, proposing therapeutic strategies to restore SNO homeostasis or target this signaling nexus for precision intervention in ER+ breast cancer (Cañas A et al., 2012).
- S- nitrosation of MBL in RA mediates immune dysfunction and autoantibody production: In patients with rheumatoid arthritis (RA), mannose-binding lectin (MBL) undergoes post-translational modification due to NO-mediated S- nitrosation, resulting in significant impairment of its function. It was found that HepG2 cells stimulated by synovial fluid in RA patients produced excessive MBL and nitric oxide, and it was confirmed by biotin switch assay that MBL was S- nitrosated. The plasma level of S- nitrosated MBL(SNO-MBL) in RA patients was significantly higher than that in healthy controls, and the mannan binding capacity, C4 deposition activity of complement, bacterial phagocytosis and apoptotic cell binding function of SNO-MBL were significantly lower than those in normal MBL. In addition, the level of autoantibodies against SNO-MBL in the plasma of RA patients increased, suggesting that this modification may change the conformation of MBL, expose potential antigenic epitopes, and thus trigger the generation of autoantibodies. These results indicate that the S- nitrosation of MBL not only directly weakens its natural immune defense function, but also may participate in the pathological process of RA by inducing autoimmune reaction, which provides a new molecular mechanism explanation for immune disorder and inflammation persistence in RA (Gupta B et al., 2008).
References
- Sun J, Hao W, Fillmore N, Ma H, Springer D, Yu ZX, Sadowska A, Garcia A, Chen R, Muniz-Medina V, Rosenthal K, Lin J, Kuruvilla D, Osbourn J, Karathanasis SK, Walker J, Murphy E. "Human Relaxin-2 Fusion Protein Treatment Prevents and Reverses Isoproterenol-Induced Hypertrophy and Fibrosis in Mouse Heart." J Am Heart Assoc. 2019 Dec 17;8(24):e013465. doi: 10.1161/JAHA.119.013465
- Cañas A, López-Sánchez LM, Peñarando J, Valverde A, Conde F, Hernández V, Fuentes E, López-Pedrera C, de la Haba-Rodríguez JR, Aranda E, Rodríguez-Ariza A. "Altered S-nitrosothiol homeostasis provides a survival advantage to breast cancer cells in HER2 tumors and reduces their sensitivity to trastuzumab." Biochim Biophys Acta. 2016 Apr;1862(4):601-610. doi: 10.1016/j.bbadis.2016.02.005
- Cañas A, López-Sánchez LM, Valverde-Estepa A, Hernández V, Fuentes E, Muñoz-Castañeda JR, López-Pedrera C, De La Haba-Rodríguez JR, Aranda E, Rodríguez-Ariza A. "Maintenance of S-nitrosothiol homeostasis plays an important role in growth suppression of estrogen receptor-positive breast tumors." Breast Cancer Res. 2012 Dec 5;14(6):R153. doi: 10.1186/bcr3366
- Zhang Y, Zhang H, Zhao S, Qi Z, He Y, Zhang X, Wu W, Yan K, Hu L, Sun S, Tang X, Zhou Q, Chen F, Gu A, Wang L, Zhang Z, Yu B, Wang D, Han Y, Xie L, Ji Y. "S-Nitrosylation of Septin2 Exacerbates Aortic Aneurysm and Dissection by Coupling the TIAM1-RAC1 Axis in Macrophages." Circulation. 2024 Jun 11;149(24):1903-1920. doi: 10.1161/CIRCULATIONAHA.123.066404
- Fujikawa K, Nakahara K, Takasugi N, Nishiya T, Ito A, Uchida K, Uehara T. "S-Nitrosylation at the active site decreases the ubiquitin-conjugating activity of ubiquitin-conjugating enzyme E2 D1 (UBE2D1), an ERAD-associated protein." Biochem Biophys Res Commun. 2020 Apr 16;524(4):910-915. doi: 10.1016/j.bbrc.2020.02.011
- Cox JB, Nukui M, Murphy EA. "Protein-S-nitrosylation of human cytomegalovirus pp65 reduces its ability to undermine cGAS." v J Virol. 2025 Apr 17:e0048125. doi: 10.1128/jvi.00481-25
- Bignon E, Allega MF, Lucchetta M, Tiberti M, Papaleo E. "Computational Structural Biology of S-nitrosylation of Cancer Targets." Front Oncol. 2018 Aug 14;8:272. doi: 10.3389/fonc.2018.00272
- Gupta B, Raghav SK, Das HR. S-nitrosylation of mannose binding lectin regulates its functional activities and the formation of autoantibody in rheumatoid arthritis." Nitric Oxide. 2008 Jun;18(4):266-73. doiI: 10.1016/j.niox.2008.01.009
- Thomas DD, Jourd'heuil D. "S-nitrosation: current concepts and new developments." Antioxid Redox Signal. 2012 Oct 1;17(7):934-6. doi: 10.1089/ars.2012.4669











