Why RiPP Genome Mining for Natural Product Discovery?
Traditional natural product discovery — bioactivity-guided screening of microbial extracts — is labor-intensive, low-throughput, and prone to rediscovering known compounds. Ribosomally synthesized and post-translationally modified peptides (RiPPs) represent a vast, underexplored reservoir of bioactive natural products encoded in microbial genomes. Genome mining offers a paradigm shift: instead of screening blindly, we computationally scan genomic data for RiPP biosynthetic gene clusters (BGCs), prioritize the most promising clusters, and experimentally validate their products through heterologous expression and high-resolution LC-MS/MS analysis. This integrated approach systematically uncovers novel RiPP scaffolds with antimicrobial, anticancer, and immunomodulatory activities that would otherwise remain silent in the genome.
What We Offer: From Genome Mining to Confirmed RiPP Structures
RiPPs are everywhere in microbial genomes — the challenge is finding the ones worth pursuing, then proving they're real. Our pipeline starts with multi-tool genome mining (antiSMASH, DeepRiPP, BAGEL4) to flag promising biosynthetic gene clusters, then moves into the lab for heterologous expression and high-resolution Orbitrap LC-MS/MS analysis. The result: a fully confirmed RiPP structure with a clean evidence trail from gene cluster to molecule.
Key Advantages of Our RiPP Genome Mining Platform
RiPP Discovery Workflow: From Genome Mining to Structural Elucidation
Our RiPP discovery workflow progresses through five well-defined stages, from microbial genome analysis to confirmed molecular structure.
Genome Sequencing & BGC Mining
BGC Prioritization & Classification
Heterologous Expression & RiPP Production
LC-MS/MS Characterization & PTM Mapping
Structural Elucidation & Reporting
1
Genome Sequencing & BGC Mining
Submit your microbial genomic DNA or assembled genome sequence. We run antiSMASH 7.0, BAGEL4, RODEO, and RiPPER to identify all RiPP BGCs and classify them by RiPP family. Output: annotated BGC map with cluster coordinates, core peptide predictions, and homology rankings.
2
BGC Prioritization & Classification
We apply prioritization filters: RRE domain presence, precursor peptide conservation, tailoring enzyme novelty, and metabolomics-guided scoring (if paired LC-MS/MS data is available). Top-ranked BGCs are selected for experimental validation. Output: prioritized BGC shortlist with rationales.
3
Heterologous Expression & RiPP Production
We clone and refactor selected BGCs into optimal expression hosts. Precursor peptide overexpression, tailoring enzyme co-expression, and media optimization are performed to maximize RiPP yield. Output: culture extracts enriched with target RiPP(s).
4
LC-MS/MS Characterization & PTM Mapping
Crude and purified RiPP fractions are analyzed by LC-MS/MS on Orbitrap Fusion Lumos using HCD, CID, and ETD fragmentation. We perform database-dependent searching, PTM-focused spectral matching, and de novo sequencing — complemented by targeted mass spectrometry quantitation for precise abundance measurements of target RiPP species. Output: annotated MS/MS spectra, PTM maps, sequence coverage.
5
Structural Elucidation & Reporting
All data are integrated into a comprehensive structural report: finalized RiPP sequence, complete PTM inventory, proposed 2D structure (with optional 3D NMR structure), BGC-to-product evidence chain, and biological activity recommendations. Output: final deliverable package.
RiPP Discovery Applications Across Therapeutic Areas
Novel Antibiotic Discovery
RiPPs include some of the most potent antimicrobial peptides known — nisin (lanthipeptide) has been used for decades as a food preservative, and lasso peptides show promising activity against multidrug-resistant Gram-negative pathogens. Genome mining systematically discovers new RiPP antibiotics with novel mechanisms of action, including lipid II binding, pore formation, and ribosomal inhibition.
Anticancer RiPP Lead Identification
Several RiPP classes — thiopeptides, borosins, and certain lanthipeptides — display potent cytotoxic activity against cancer cell lines. Our genome mining pipeline prioritizes RiPP BGCs with predicted bioactivities relevant to oncology, enabling the discovery of new peptide-based anticancer leads.
Microbiome-Derived RiPP Exploration
Human and environmental microbiomes harbor an immense diversity of RiPP BGCs that remain functionally uncharacterized. Metagenome mining of microbiome datasets reveals RiPP clusters from uncultured microbial lineages, opening access to truly novel RiPP chemotypes.
Enzyme Engineering of RiPP Biosynthetic Machinery
The tailoring enzymes that install RiPP PTMs — lanthionine cyclases, dehydratases, methyltransferases, P450s — are themselves valuable biocatalysts. Characterizing RiPP pathways from genome mining enables enzyme discovery and re-engineering for applications in peptide macrocyclization and bioorthogonal modification.
RiPP-Based Diagnostic Probe Development
RiPPs with specific receptor-binding activities, such as lasso peptide receptor antagonists and microviridin-type enzyme inhibitors, can be developed as molecular probes or diagnostic leads. Genome mining identifies RiPP candidates with predictable target-binding modalities for diagnostic applications.
RiPP Discovery Platform: Representative Results
Representative data from our RiPP genome mining and structural analysis platform demonstrate the quality and depth of information we deliver at each stage of the discovery pipeline, complementing our untargeted peptidomics profiling workflows for comprehensive peptide natural product characterization.
BGC Genomic Architecture Map
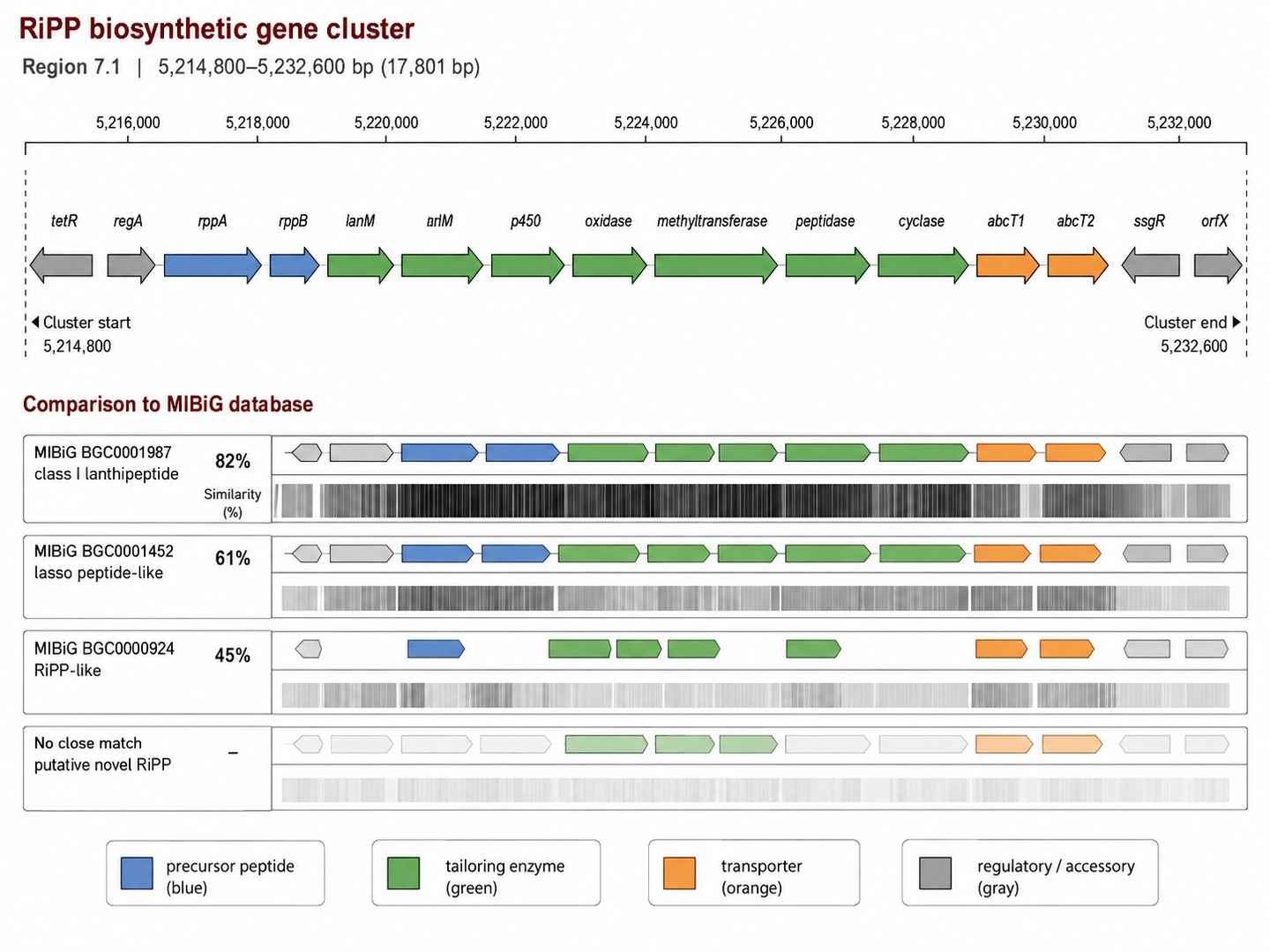
AntiSMASH 7.0 visualization of a RiPP BGC identified from a Streptomyces genome, showing core biosynthetic genes, regulatory elements, and cluster boundary predictions.
LC-MS/MS Characterization of RiPP Product
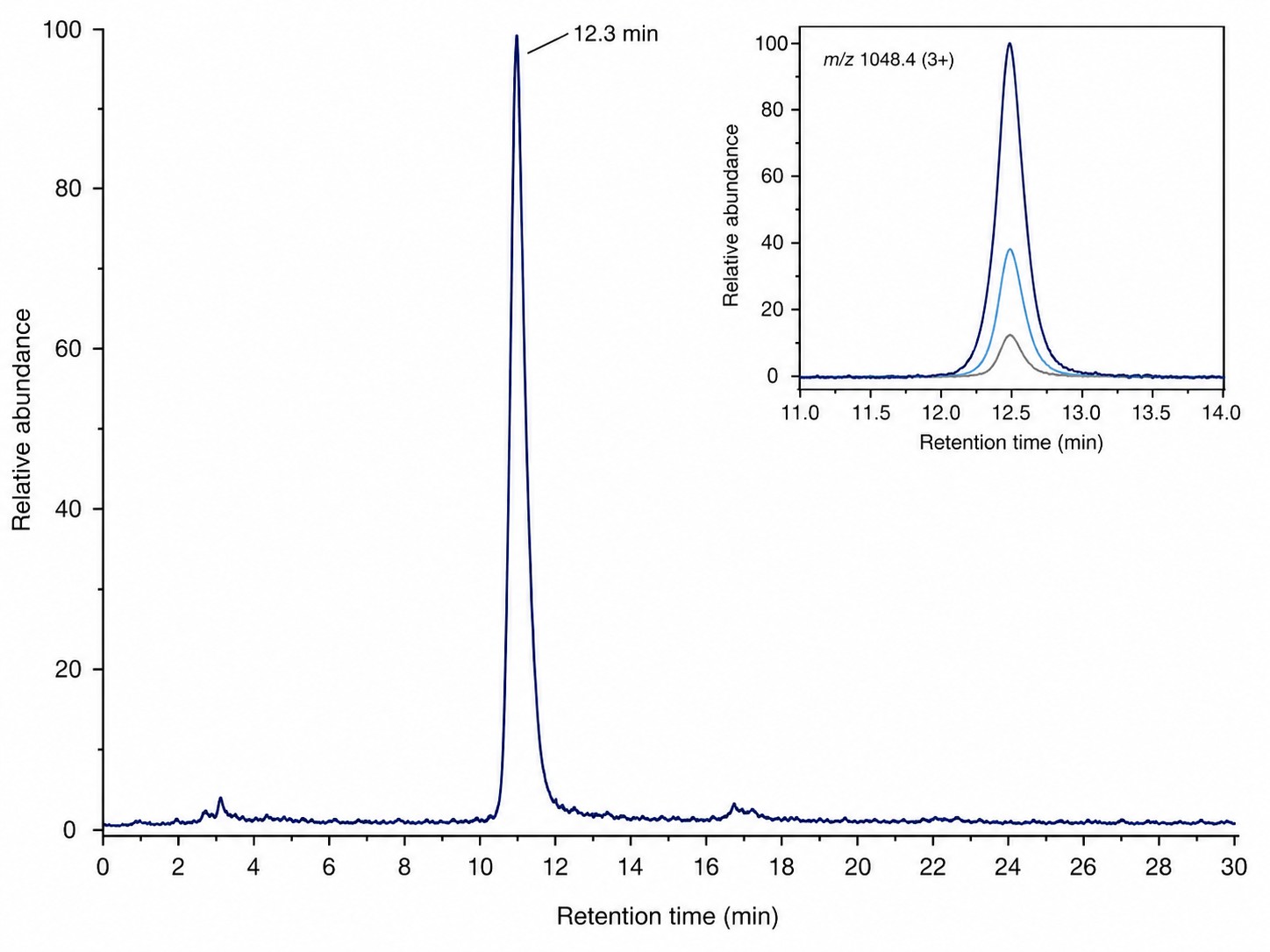
Base peak chromatogram and EICs from Orbitrap Fusion Lumos analysis confirming the target RiPP product with baseline resolution and sub-2 ppm mass accuracy.
PTM Mapping — Lanthionine Bridge Topology
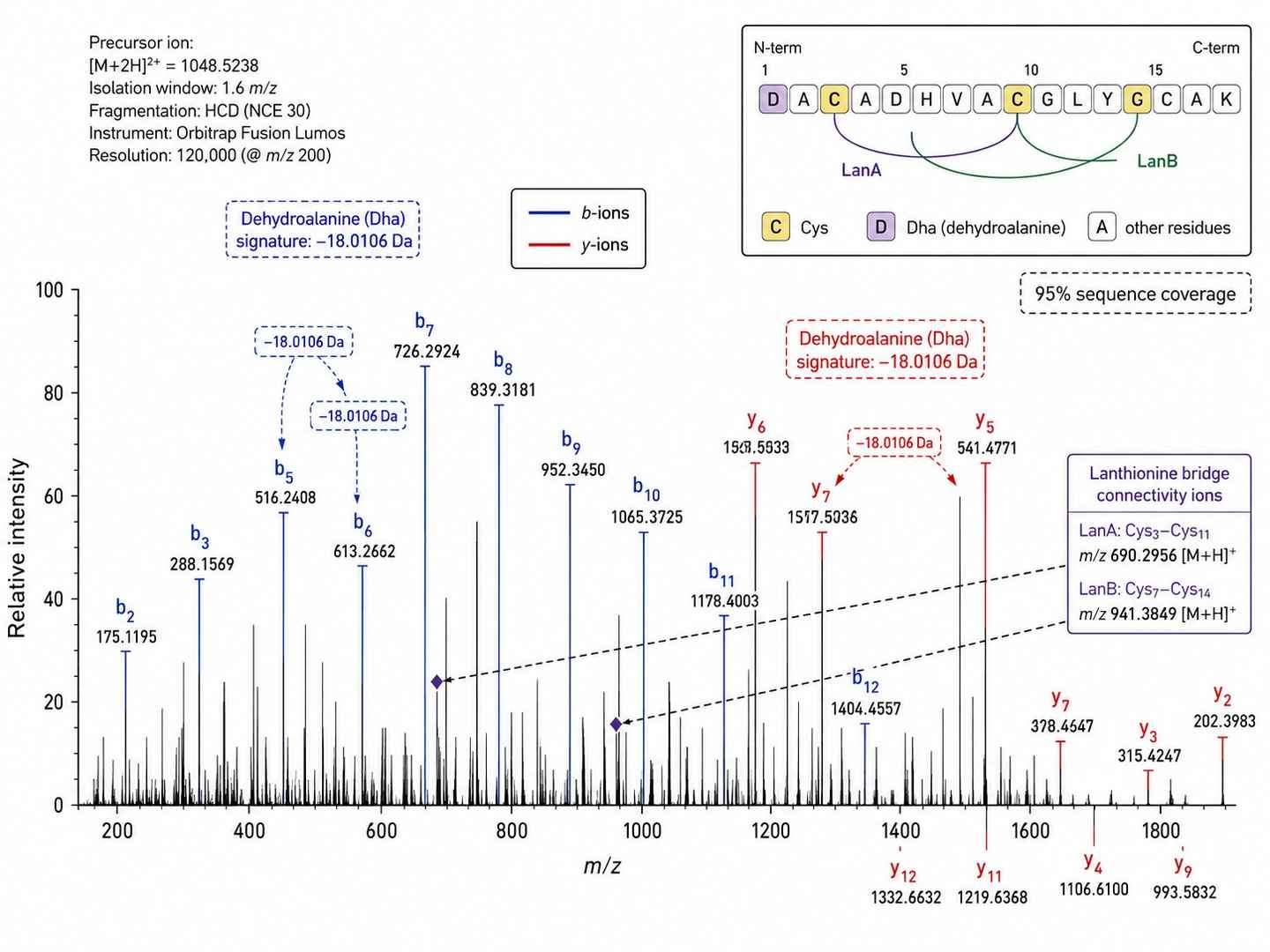
Annotated HCD MS/MS spectrum showing 95% sequence coverage with unambiguous assignment of lanthionine bridge connectivity and dehydroalanine signatures.
3D Structural Model of Characterized RiPP
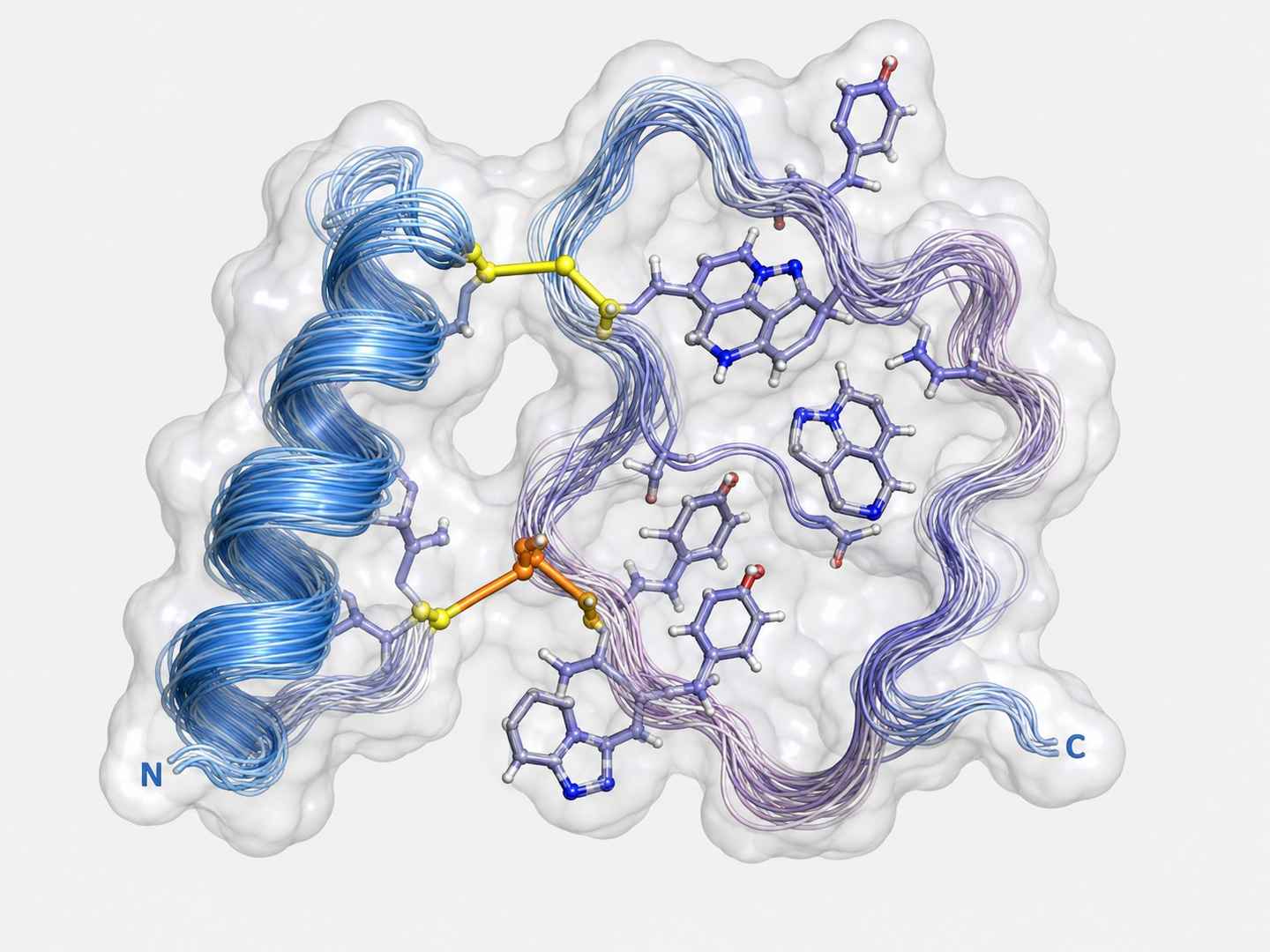
Solution NMR-derived (600 MHz, 2D NOESY/TOCSY) structural ensemble showing lanthionine bridge cross-links and spatial arrangement of key residues.
Sample Requirements for RiPP Genome Mining Projects
To ensure successful RiPP discovery and characterization, appropriate sample quality and quantity are essential. Below are our standard requirements for the most common sample types.
| Sample Type | Quantity Required | Quality Requirements | Shipping Conditions |
|---|---|---|---|
| Genomic DNA | ≥ 1 μg | OD260/280: 1.8–2.0; OD260/230: ≥ 1.5; no visible degradation | DNase-free tube; ship on dry ice or ice packs |
| Purified RiPP | ≥ 10 pmol (≥ 50 pmol for NMR) | > 80% purity by HPLC-UV; MS-compatible buffer | Shipped on dry ice; avoid freeze-thaw cycles |
| Bacterial Strain | Live culture or glycerol stock | Verified genotype; axenic culture; antibiotic markers specified | Room temperature (live plate) or −80 °C (glycerol stock) |
| Crude Extract | ≥ 100 μL or ≥ 10 mg dry weight | Organic extract (MeOH, EtOAc, or ACN/H2O) | Shipped on dry ice; protect from light |
For metagenomic samples (soil, gut, marine), please consult our scientific team for minimum DNA yield and sequencing depth requirements. A minimum of 5 Gb of shotgun metagenomic sequence data is recommended for comprehensive RiPP BGC mining from complex microbial communities.
RiPP Discovery Project Deliverables
We deliver a comprehensive data package that provides a complete evidence chain from genome mining prediction to confirmed RiPP structure.
- Annotated BGC Map
Genome browser-compatible visualization (GBK, GFF) of all identified RiPP BGCs with gene annotations, precursor sequences, domain architectures, and antiSMASH scores. - RiPP Sequence & PTM Profile
Confirmed full-length sequence with single-amino-acid resolution. Complete PTM inventory including modification types, positions, and bridge connectivity maps. - LC-MS/MS Raw Data
Full Orbitrap data package: .raw files, peak lists (.mgf, .mzML), and annotated spectra in PDF and interactive HTML formats. - Structural Model Report
2D/3D structure with ensemble statistics and PDB coordinates (NMR) or per-residue confidence scoring (MS-only). - De Novo Sequencing Results
PEAKS Studio output with average local confidence scores per residue, ranked candidates, and expert-reviewed validation notes. - Comprehensive Final Report
Integrated discovery narrative from genome mining predictions to final RiPP structure, including QC metrics and bioactivity testing recommendations.
Frequently Asked Questions (FAQ)
What is RiPP genome mining and how does it accelerate natural product discovery? +
RiPP genome mining is a computational approach that systematically scans microbial genome sequences for biosynthetic gene clusters encoding RiPPs. Unlike traditional bioactivity-guided screening — which can take months to years and repeatedly rediscovers known compounds — genome mining can process thousands of genomes in silico within days, prioritize BGCs encoding structurally novel RiPPs, and guide targeted experimental validation. This approach has accelerated the discovery rate of new RiPP classes from years per class to multiple classes per year, as demonstrated by the identification of imiditides, two-amino-terminus RiPPs, and sulfonated lanthipeptides within the past three years.
Which RiPP classes can you detect and characterize? +
Our pipeline covers all major RiPP classes recognized by the MiBIG and antiSMASH ontologies, including lanthipeptides (classes I–IV), lasso peptides, thiopeptides, graspetides, linariidins, proteusins, sactipeptides, head-to-tail cyclized peptides, glycocin-type RiPPs, borosins, and RaS-RiPPs. We also maintain validated protocols for emerging classes such as imiditides, two-amino-terminus RiPPs, and thioamidated RiPPs. For BGCs that do not match known classes, our de novo characterization approach is designed precisely for such cases.
How do you validate antiSMASH BGC predictions experimentally? +
Our validation workflow proceeds through three stages: (1) heterologous expression — we clone and refactor the predicted BGC into a suitable host and optimize expression conditions; (2) targeted LC-MS/MS analysis — we monitor for ions matching the predicted core peptide mass within 3 ppm accuracy, then fragment them using HCD/ETD to confirm sequence and PTMs; (3) structural confirmation — we compare experimental MS/MS spectra against predicted fragmentation patterns, apply de novo sequencing, and for high-value targets, perform NMR-based 3D structure determination. This three-stage approach achieves over 80% success rate for well-prioritized BGCs.
What mass spectrometry platforms do you use for RiPP characterization? +
We operate Orbitrap Fusion Lumos (Thermo Fisher) and timsTOF Pro (Bruker) platforms. The Orbitrap delivers ultra-high resolution (up to 500,000 at m/z 200) and multi-stage fragmentation — critical for disambiguating isobaric PTM variants. The timsTOF Pro adds ion mobility separation (TIMS) to resolve RiPP conformers and isomeric macrocyclic topologies. Both platforms are coupled to nanoLC and microLC systems optimized for low femtomole sensitivity.
Can you characterize RiPPs from unusual sample types (microbiome, environmental samples)? +
Yes. For metagenomic samples, we accept shotgun metagenomic sequencing data and perform assembly, binning, and RiPP BGC mining from metagenome-assembled genomes. We also offer metabolomics-guided detection comparing LC-MS/MS data against in silico predicted RiPP masses. This approach effectively connects RiPP BGCs in metagenomes to their actual products without requiring culture-based isolation. For human microbiome samples, we perform strain-level RiPP BGC mining and prevalence analysis across disease versus healthy populations.
Disclaimer: All services and analytical platforms described are intended for translational research and preclinical support. Research Use Only (RUO). Not for use in diagnostic procedures.
Case Study
Discovery of Two-Amino-Terminus RiPPs via RRE-Guided Genome Mining
Journal: Nature Communications
Published: 2023
Summary
In this landmark study, Ren et al. (University of Illinois Urbana-Champaign) used RRE-guided genome mining to discover a previously unknown RiPP class characterized by two functional amino termini, named "mpa" RiPPs. The integrated workflow — combining RRE-Finder genome scanning, heterologous expression in E. coli, and Orbitrap Fusion Lumos LC-MS/MS with HCD/ETD fragmentation — successfully characterized the 28-amino-acid MpaA RiPP, revealing a unique dual-cleavage biosynthetic logic that yields an unprecedented dual-amino-terminus architecture.
Methods
The research team employed RRE-Finder to scan microbial genome databases for RiPP recognition element domains. Using RRE sequences from characterized lanthipeptide systems as baits, they identified over 200 candidate RiPP BGCs across Actinobacteria genomes. One candidate cluster (mpa) from Actinoplanes missouriensis was prioritized for its unusual architecture — encoding a precursor peptide with two RiPP core motifs flanking a central RRE domain. The mpa BGC was refactored and heterologously expressed in E. coli BL21(DE3), and culture extracts were analyzed by LC-MS/MS on an Orbitrap Fusion Lumos using HCD and ETD fragmentation, with 2D NMR (NOESY, TOCSY) for final structural confirmation.
Results
LC-MS/MS analysis confirmed the expression of a 28-amino-acid RiPP product (MpaA, 3,142.5 Da) with 96% sequence coverage by ETD fragmentation. Annotated MS/MS spectra revealed a unique dual cleavage event — processing at both N-terminal and internal recognition sites — yielding a mature RiPP with two free amino termini, an architecture never before reported in any RiPP class. Further characterization demonstrated that the RRE domain recruits two distinct modification enzymes: MpaB (lanthionine cyclase) and MpaC (dehydratase), which install three dehydroalanine residues and one lanthionine bridge (DL-configuration confirmed by 2D NMR). This integrated genome mining and structural analysis approach demonstrates that even deeply hidden RiPP classes defined by unprecedented PTM topology are discoverable through a well-executed pipeline.
References
- Ren H, Dommaraju SR, Huang C, et al. Genome mining unveils a class of ribosomal peptides with two amino termini. Nature Communications. 2023;14:1624. doi:10.1038/s41467-023-37287-1
- Blin K, Shaw S, Augustijn HE, et al. antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Research. 2023;51(W1):W46-W50. doi:10.1093/nar/gkad344
- Skinnider MA, Johnston CW, Edgar RE, et al. Genomic charting of ribosomally synthesized natural product chemical space facilitates targeted discovery. PNAS. 2016;113(42):E6343-E6351. doi:10.1073/pnas.1609014113
- van Heel AJ, de Jong A, Song C, et al. BAGEL4: a user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Research. 2018;46(W1):W278-W281. doi:10.1093/nar/gky383
- Tietz JI, Schwalen CJ, Patel PS, et al. A new genome-mining tool redefines the lasso peptide biosynthetic landscape. Nature Chemical Biology. 2017;13:470-478. doi:10.1038/nchembio.2318
- Merwin NJ, Mousa WK, Dejong CA, et al. DeepRiPP integrates multiomics data to automate discovery of novel ribosomally synthesized natural products. PNAS. 2020;117(1):371-380. doi:10.1073/pnas.1901493116