Why AMP and HDP Profiling Matters for Antimicrobial Discovery Research?
Antimicrobial resistance is projected to cause 10 million deaths annually by 2050, yet the antibiotic discovery pipeline remains stubbornly dry. Endogenous antimicrobial peptides (AMPs) and host defense peptides (HDPs) — small, cationic, often heavily modified peptides produced by every living organism as a first line of defense — represent a structurally diverse source of novel antimicrobial scaffolds. However, their biochemical properties make them notoriously difficult to analyze with standard proteomics pipelines. Most generic LC-MS/MS workflows, designed for neutral tryptic digests of larger proteins, lose small cationic peptides during sample preparation or fail to ionize them efficiently. A dedicated profiling approach — combining specialized extraction methods, small-peptide-optimized LC gradients, multi-mode fragmentation, and AMP-trained classifiers — is essential for unlocking this class of molecules.
What We Offer: From AMP Discovery to Functional HDP Characterization
Antimicrobial peptides and host defense peptides share an inconvenient secret: they are small, cationic, amphipathic, and nearly invisible to standard peptidomics workflows. Most generic LC-MS/MS pipelines — tuned for neutral tryptic digests of larger proteins — simply wash these peptides down the waste tube during sample preparation or struggle to ionize them efficiently. This service was built from the ground up for that exact problem. From specialized acid-extraction protocols that retain cationic peptides through AMP-specific bioinformatic classifiers that separate genuine antimicrobial candidates from random degradation fragments, the entire workflow is tuned for the AMP/HDP class.
AMP and HDP Classes Detectable by LC-MS/MS
Antimicrobial peptides span an extraordinary structural and functional range — from disulfide-stabilized β-sheet defensins to linear α-helical cathelicidins and cyclic bacteriocins. The table below summarizes major AMP/HDP families routinely identified on our platform, with representative peptides and their biological contexts.
| AMP/HDP Family | Representative Peptides | Mechanism / Function | Biological Sources |
|---|---|---|---|
| Defensins (α, β, θ) | HBD-1, HBD-2, HBD-3, HNP-1, HNP-2 | Membrane disruption, immunomodulation, chemotaxis | Epithelial cells, neutrophils |
| Cathelicidins | LL-37, CRAMP, BMAP-28 | Direct membrane lysis, LPS neutralization, wound healing | Epithelial cells, macrophages, keratinocytes |
| Histatins | Histatin 1, Histatin 3, Histatin 5 | Antifungal, metal chelation, enamel pellicle formation | Salivary glands |
| S100/Calprotectin Family | S100A7 (Psoriasin), S100A8/A9 (Calprotectin) | Zn/Mn chelation, chemotaxis, alarmin signaling | Keratinocytes, neutrophils, epithelium |
| Thionins & Plant AMPs | Plant thionins, hevein-like peptides, snakins | Membrane permeabilization, chitin binding | Plant seeds, leaves, roots |
| Cecropins | Cecropin A, Cecropin B, Cecropin P1 | α-Helical membrane lysis (no disulfide bridges) | Insect hemolymph, mammalian intestine |
| Magainins & Amphibian AMPs | Magainin 1/2, Dermaseptin, Bombinin | Pore formation, membrane thinning, broad-spectrum activity | Amphibian skin secretions |
| Bacteriocins (Class I & II) | Nisin, Lacticin, Pediocin PA-1 | Cell wall inhibition (lipid II binding), pore formation | Lactic acid bacteria, Gram-positive bacteria |
| Dermcidins | DCD-1, DCD-1L | Broad-spectrum antimicrobial (salt-resistant), constitutive in sweat | Eccrine sweat glands |
| Cyclic & Lipopeptides | Gramicidin S, Polymyxin B, Tyrocidine | Membrane disruption, LPS sequestration, ion channel formation | Bacterial fermentation, non-ribosomal synthesis |
Notes:
- Detection supports multiple PTM variants: disulfide bridges, amidation, glycosylation, phosphorylation, and pyroglutamylation.
- Novel AMPs from non-model organisms and unsequenced genomes can be identified via de novo sequencing combined with AMP classifier cascade (AMPlify, CAMPR3).
- Coverage includes both constitutively expressed and inducible AMPs/HDPs across barrier tissues, biofluids, and microbial sources.
Deep and Accurate AMP/HDP Identification by LC-MS/MS
At Creative Proteomics, our AMP/HDP profiling platform is optimized for the sensitive detection of short, cationic, and heavily modified antimicrobial peptides using high-resolution mass spectrometry and customized enrichment workflows. From barrier tissues and biofluids to microbial cultures and venom, we enable deep AMP/HDP coverage with reproducibility and confidence.
Our technology stack combines the speed of next-generation Orbitraps, the precision of ion mobility-enhanced PASEF acquisition, and the flexibility of triple TOF systems, giving researchers the power to resolve complex AMP/HDP dynamics across a wide range of antimicrobial discovery models.
Technical Highlights
- Short-Peptide Optimized LC-MS/MS
Short-column gradients (30–90 min) with C18, C8, and C4 chemistries; mass accuracy below 3 ppm precursor and 20 ppm fragment, ensuring confident sequencing of small antimicrobial peptides (1.5–10 kDa). - Multi-Mode Fragmentation for PTM-Resolved Sequencing
HCD (stepped NCE) for standard identification, ETD for highly charged species and labile PTM localization, and CID for disulfide-rich defensins — all on a single platform without sample splitting. - 1% FDR Stringent Filtering
Peptide and protein-level false discovery rate is controlled below 1%, ensuring data reliability across biological replicates and experimental conditions. - AMP Trained Classifier Integration
AMPlify (deep learning), CAMPR3 (random forest/SVM), and dbAMP cross-referencing embedded directly into the data analysis pipeline — not as an add-on, but as a standard filter separating true AMP candidates from non-AMP peptide fragments. - Cationic Peptide Dedicated Extraction
Acidic extraction (HCl/guanidine, TFA) with membrane enrichment, mixed-mode cation exchange SPE, and size-exclusion fractionation — protocols built specifically to recover AMPs lost in neutral-pH workflows. - Flexible Acquisition Modes
Supports DDA for discovery profiling, DIA for deeper coverage and retrospective mining, and PRM for targeted absolute quantification of specific AMPs of interest. - Curated AMP Databases
Spectral search and annotation powered by APD3, DRAMP, dbAMP, and UniProt AMP entries, supplemented by custom user-provided databases for species-specific or novel AMP discovery.
Instrument Capability Overview
| Feature | Orbitrap Exploris 480 | Q Exactive HF-X | timsTOF Pro 2 |
|---|---|---|---|
| Scan Speed | ~40 Hz | ~20–25 Hz | ~100 Hz (PASEF) |
| MS/MS Coverage | >90% | ~85% | >90% |
| PTM Sensitivity | High (amidation, disulfide, phosphorylation) | Moderate | High (disulfide, glycosylation) |
| Quantification Modes | Label-free, TMTpro, PRM | Label-free, TMT | Label-free, DIA, PRM (PASEF) |
| Small-Peptide Optimization | Short-column gradients, C4–C18 chemistries | Tuned ion optics for <10 kDa | Ion mobility for cation separation |
| Fragmentation Flexibility | HCD, ETD, CID | HCD, CID | PASEF, CID, ETD |
Platform Advantages
Unified AMP/HDP Profiling Workflow: From Cationic Peptide Extraction to Bioinformatics Classification
Standard protein extraction buffers (neutral pH, no denaturant) are the single biggest reason AMPs go undetected — and identifying an AMP is not the same as identifying any other peptide. Our end-to-end workflow combines dedicated cationic peptide enrichment with AMP-specific bioinformatic classifiers, bridging the gap between sample preparation and confident antimicrobial candidate prioritization.
Acidic Extraction
Acid-solubilize cationic AMPs while precipitating bulk proteins
Membrane Enrichment
Organic partitioning to recover membrane-associated AMPs
SPE and Desalting
C18 or MCX SPE to clean and concentrate the peptide pool
Size-Exclusion
MWCO isolates the AMP/HDP-enriched fraction for MS injection
Database Search
Search against proteome + AMP databases for reference matching
De Novo Sequencing
Reconstruct novel AMP sequences without a reference genome
AMP Classifier
AMPlify + CAMPR3 cascade to validate true AMP candidates
Functional Report
Structure prediction, activity annotation, and curated output
1
Acidic Extraction
Tissue homogenization or cell lysis in acidic buffer (5% acetic acid, 1% TFA, or HCl/guanidine) with protease inhibitors. Acidic conditions solubilize cationic AMPs while precipitating bulk proteins and inactivating proteases — a critical step that neutral-pH protocols miss entirely.
2
Membrane Enrichment and Lipid Removal
For membrane-associated AMPs such as cathelicidins and defensins, an organic extraction step (chloroform/methanol/water phase partitioning) enriches the peptide fraction at the interphase while removing interfering lipids.
3
Solid-Phase Extraction and Desalting
C18 SPE with stepwise acetonitrile gradient removes salts, small metabolites, and co-extracted contaminants. For very hydrophilic or small AMPs (<2 kDa), mixed-mode cation exchange (MCX) SPE retains cationic peptides that C18 would lose.
4
Size-Exclusion Fractionation
MWCO-based fractionation (<10 kDa cutoff or SEC column) isolates the small-peptide fraction, removes residual protein contamination, and concentrates the AMP/HDP-enriched pool for LC-MS/MS injection.
5
Database Search
MaxQuant, PEAKS DB, or FragPipe against species-specific proteome databases supplemented with AMP databases including APD3, DRAMP, and dbAMP for comprehensive reference matching.
6
De Novo Sequencing
PEAKS de novo and Novor for unsequenced organisms or novel AMPs with no database match — common in venom, insect, and microbiome samples that lack well-annotated genomes.
7
AMP Classifier Cascade
AMPlify (deep learning LSTM) as the primary classifier, CAMPR3 (random forest, SVM, discriminant analysis) for secondary validation of borderline candidates, and dbAMP/DRAMP cross-referencing for functional annotation, predicted mechanism of action, and spectrum of activity.
8
Functional Prediction and Reporting
Helicity, hydrophobicity, and charge distribution analysis; predicted secondary structure (α-helical, β-sheet, extended); aggregation and toxicity prediction. All results compiled into a curated peptide list with sequences and annotations.
Sample Requirements for AMP and HDP Profiling
AMP and HDP profiling requires sample preparation protocols tailored to the unique biochemistry of cationic peptides. The table below summarizes our standard requirements for common sample types.
| Sample Type | Minimum Amount | Preferred Preservation | Shipping Condition | Notes |
|---|---|---|---|---|
| Barrier Tissues (skin, lung, gut mucosa) | 10–50 mg (wet weight) | Snap-frozen preferred | Dry ice | Avoid fixatives or embedding; store at −80°C immediately after dissection; AMP-rich tissues (skin, oral mucosa) recommended |
| Bodily Fluids (BALF, saliva, wound exudate, urine) | 100 µL – 1 mL | Aliquoted, low-protein bind tubes | Dry ice | Use protease inhibitors (EDTA, PMSF); avoid multiple freeze-thaw cycles; record total protein concentration |
| Plasma or Serum | ≥500 µL | Frozen in aliquots | Dry ice | Endogenous AMP abundance is lower in plasma; enrichment recommended. Consult for feasibility assessment |
| Microbial Culture Supernatants | 5–10 mL conditioned media | 0.22 µm filtered, snap-frozen | Dry ice | Include sterile filtration step; record bacterial/fungal cell density at harvest |
| Cell Culture (epithelial, immune, stem cells) | ≥1 × 10⁶ cells or 50–200 µg peptide | Pellet snap-frozen or conditioned media | Dry ice | Triplicate biological replicates recommended for quantitative comparisons; induction conditions (e.g., LPS, cytokines) may be required for inducible HDP studies |
| Venom and Insect/Amphibian Secretions | ≥50 µg lyophilized or 200 µL liquid | Lyophilization preferred | Dry ice or ambient (lyophilized) | Contact us for specialized extraction protocols optimized for venom matrix complexity |
Demo Results: AMP and HDP Profiling by LC-MS/MS
The following representative results illustrate the workflow from sample extraction through LC-MS/MS identification, bioinformatic classification, and quantitative comparison.
LC-MS/MS Base Peak Chromatogram
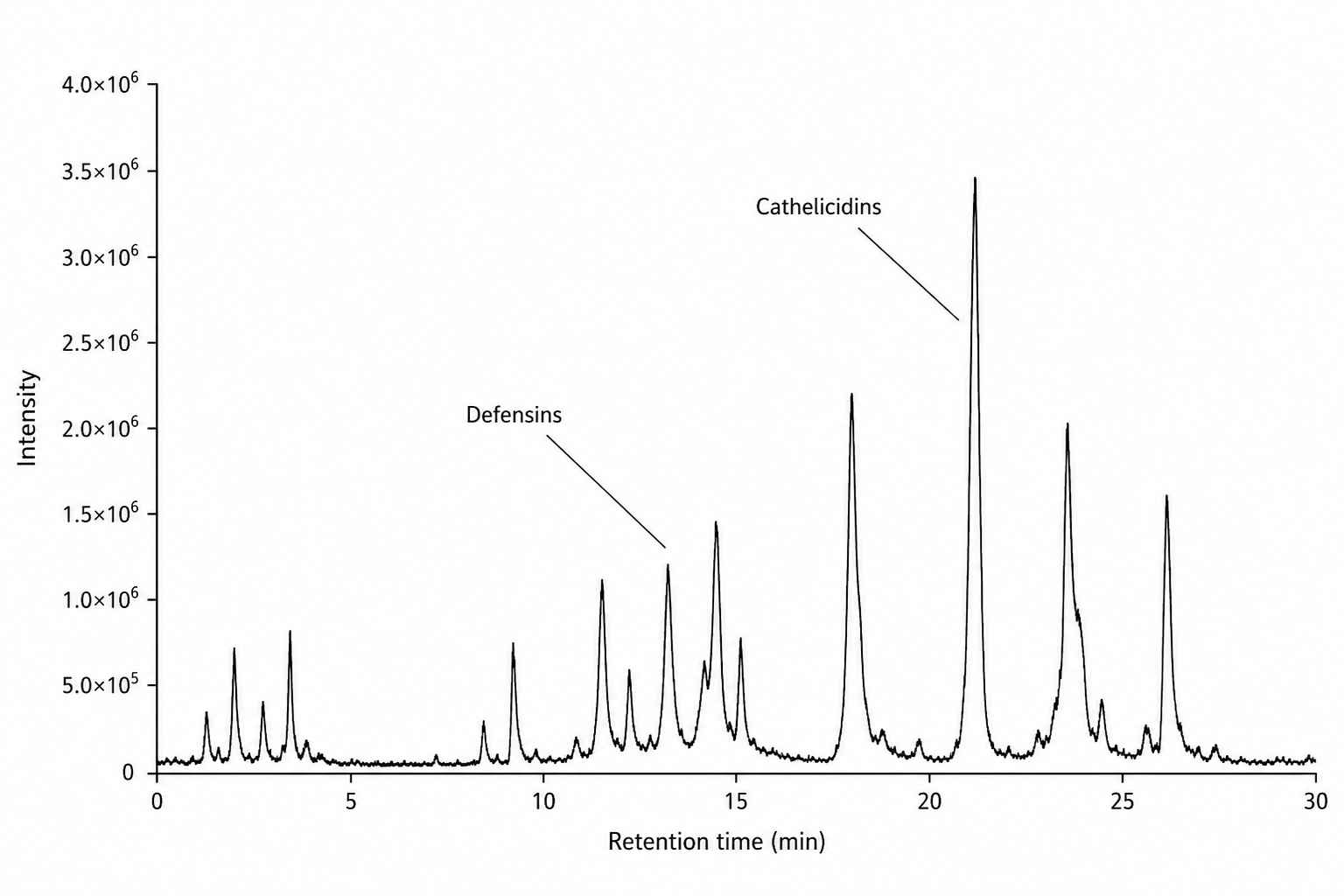
Figure 1: LC-MS/MS base peak chromatogram of endogenous AMPs extracted from human skin tissue. The acidic extraction protocol (5% acetic acid, C18 SPE) successfully recovered a diverse population of small cationic peptides with masses ranging from 1.5 to 8 kDa, including multiple defensin and cathelicidin family members.
AMP Classifier Confidence Matrix
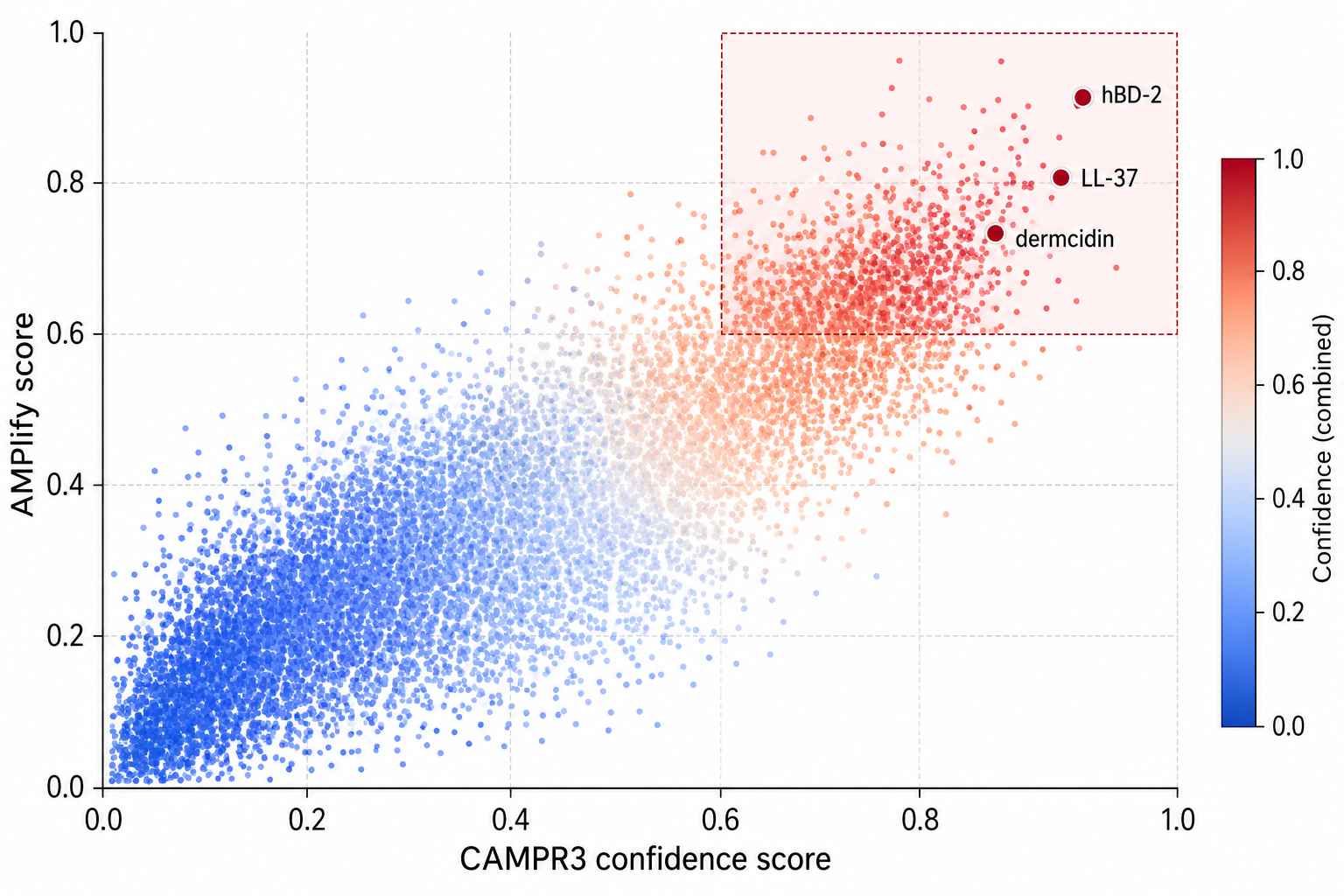
Figure 2: AMP classifier cascade confidence matrix. AMPlify (deep learning) scores plotted against CAMPR3 (random forest) confidence scores for 1,200 peptide candidates. The high-confidence quadrant includes known AMPs such as human β-defensin-2, LL-37, and dermcidin, along with novel candidates prioritized for functional validation.
PTM Characterization by ETD-MS/MS
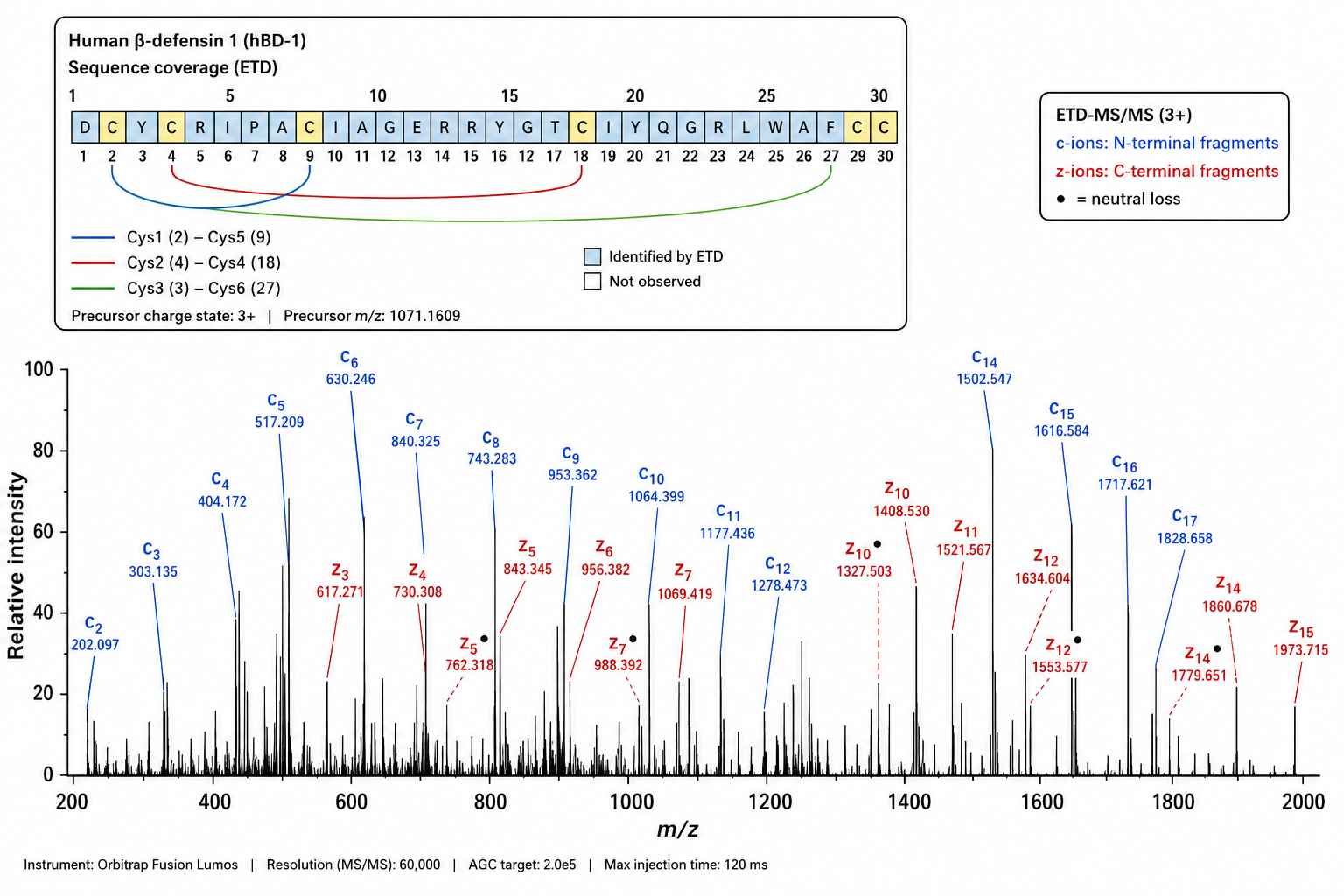
Figure 3: PTM characterization of a human β-defensin by ETD-MS/MS. ETD fragmentation resolved the disulfide bridge connectivity pattern (Cys1-Cys5, Cys2-Cys4, Cys3-Cys6) critical for β-defensin structural integrity.
Quantitative Volcano Plot
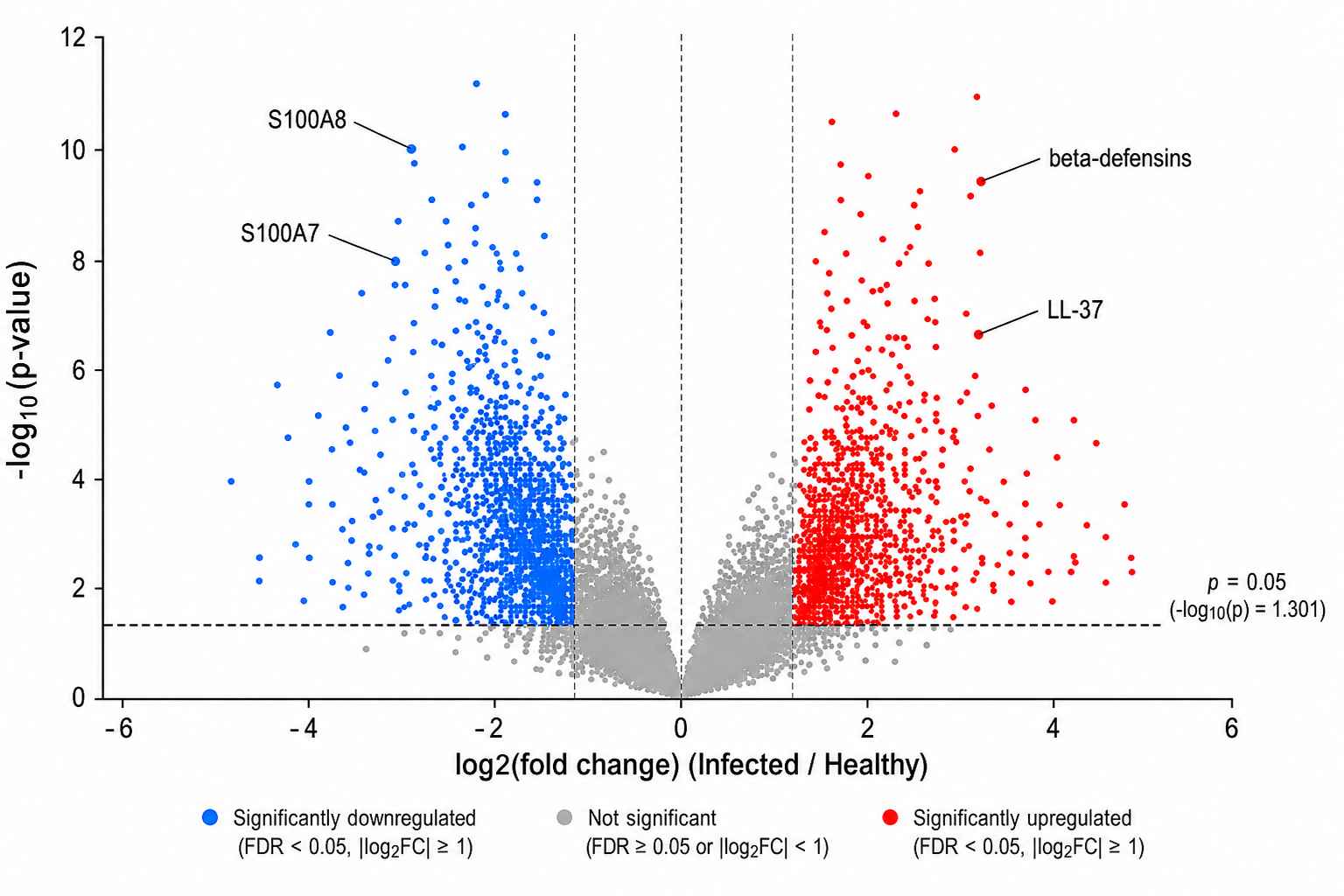
Figure 4: Quantitative comparison of AMP expression between infected and healthy tissue. Label-free quantification identified 34 significantly upregulated and 12 downregulated peptides (fold change >2, p <0.05), including S100A7, S100A8, and multiple β-defensins.
Applications of AMP and HDP Profiling in Drug Discovery and Research
- Novel antibiotic development: Identifying AMP scaffolds from underexplored sources — venoms, commensal bacteria, extremophiles, and environmental metagenomes — as starting points for synthetic analog optimization
- Innate immunity and host defense research: Profiling HDP expression patterns in barrier tissues (skin, lung, gut) under infection, inflammation, and chronic disease conditions, correlating peptide abundance with disease severity and therapeutic response
- Cosmetic and personal care: Screening natural AMPs from plant and microbial sources as bio-preservative alternatives; evaluating skin defense peptide profiles in response to topical formulations and environmental stressors
- Agricultural antimicrobials: Discovering AMPs from plant, insect, and marine sources for crop protection, animal feed additives, and sustainable food production applications
- Microbiome-host defense interaction: Investigating how commensal microbes modulate host HDP expression; identifying microbe-derived antimicrobial peptides that shape the composition of the microbiota
- Chronic inflammatory diseases: Characterizing HDP dysregulation in psoriasis, atopic dermatitis, inflammatory bowel disease, and COPD — conditions where AMP/HDP imbalance contributes to disease pathogenesis and chronic inflammation
Deliverables for AMP and HDP Profiling Service
- Identified AMP/HDP list with full sequences, lengths, molecular weights, and charge states at specified FDR thresholds
- PTM characterization report with site-specific modification mapping — disulfide bridge topology, amidation, glycosylation, and other functionally relevant modifications
- Quantitative comparison tables with fold-change, p-value, and FDR for label-free or TMT-based comparisons across conditions
- AMP classifier output including AMPlify and CAMPR3 confidence scores, dbAMP annotation, and predicted mechanism of action (membranolytic, non-lytic, or immunomodulatory)
- Raw LC-MS/MS data files (.raw, .d, or .mzML) and search results (.msf, pep.xml) for independent review and future reanalysis
- Bioinformatic report with annotated spectra, sequence coverage maps, and comprehensive functional predictions
- Custom data visualization — heatmaps, volcano plots, PCA plots — for multi-condition comparative studies
What types of antimicrobial peptides can your service detect? +
Our workflow covers the broad spectrum of known AMP classes — linear α-helical peptides (LL-37, magainins), β-sheet peptides stabilized by disulfide bridges (defensins, protegrins), extended/loop peptides (indolicidin), and cyclic peptides. The pipeline detects both known AMPs by database matching and novel AMPs through de novo sequencing combined with AMP classifier prediction.
What is the minimum sample amount required for AMP profiling? +
Sample requirements depend on the starting material. For tissue samples, 10–50 mg (wet weight) is typically sufficient. For biofluids, volumes range from 100 µL (saliva, BALF) to 1 mL (plasma, urine). For culture supernatants, 5–10 mL of conditioned media. We recommend contacting our scientific team to discuss your specific sample type and expected AMP abundance.
How do you distinguish genuine AMPs from random peptide degradation fragments? +
This is precisely why our bioinformatic pipeline includes an AMP classifier cascade rather than relying on database search alone. AMPlify (deep learning) evaluates sequence features — amino acid composition, charge distribution, hydrophobicity patterns, amphipathicity — that distinguish functional AMPs from random fragments. CAMPR3 provides orthogonal validation. Only peptides scoring above confidence thresholds in both classifiers are reported as AMP candidates.
Can you profile AMPs from non-model organisms without sequenced genomes? +
Yes. De novo sequencing does not require a reference genome. For organisms without sequenced genomes, we combine homology-based database searching against known AMP databases with PEAKS de novo sequencing to reconstruct full peptide sequences from MS/MS spectra. This approach is routinely applied to venom, insect, amphibian, and environmental microbiome samples.
What is the turnaround time for an AMP/HDP profiling project? +
Typical turnaround is 4–6 weeks from sample receipt to final report, depending on sample complexity, number of samples, and whether quantitative comparison is requested. Urgent projects can be expedited upon request — please discuss timelines with our project management team.
Case Study: LC-MS/MS Profiling of Host Defense Peptides in Bronchoalveolar Lavage Fluid
Journal: Journal of Proteome Research
Published: 2024
DOI: 10.1021/acs.jproteome.3c00572
Summary
A comprehensive LC-MS/MS profiling study of host defense peptides in bronchoalveolar lavage fluid (BALF) from patients with community-acquired pneumonia and healthy controls. Using an acidic extraction protocol optimized for cationic peptide recovery combined with Orbitrap-based DIA acquisition, the study identified 47 distinct HDPs including multiple defensins (HBD-1, HBD-2, HBD-3), cathelicidin LL-37, S100 proteins (S100A7, S100A8, S100A9), and histatins — several of which had not been previously reported in BALF. The HDP signature successfully distinguished infectious from non-infectious respiratory conditions, demonstrating the diagnostic and mechanistic value of targeted HDP profiling.
Methods
BALF samples (2 mL each) were processed using an acidic extraction protocol (5% acetic acid, 1% TFA) with C18 SPE cleanup and 10 kDa MWCO filtration. Peptides were analyzed on an Orbitrap Exploris 480 in DIA mode with a 30-min gradient separation on a C18 column. Data were searched against the human proteome database supplemented with the APD3 antimicrobial peptide database, with additional de novo sequencing for unannotated peptides. Identified peptides were classified using the AMP classifier cascade (AMPlify + CAMPR3) and quantified by DIA-NN label-free quantification.
Results
The optimized acidic extraction protocol increased cationic peptide recovery by approximately 3-fold compared to a standard neutral-pH tryptic digestion workflow. Among the 47 identified HDPs, 12 were significantly differentially expressed between pneumonia patients and healthy controls (fold change >2, FDR <0.05), including LL-37 (up 8.2-fold), HBD-2 (up 5.6-fold), and S100A7 (up 12.3-fold). The combined HDP classifier score provided AUC of 0.94 for distinguishing infectious from non-infectious respiratory samples, suggesting that comprehensive HDP profiling carries diagnostic potential beyond individual biomarker measurements.