
Key takeaways
- Brain- and neuron-derived extracellular vesicles (BDEs/NDEVs) provide a blood-accessible readout of CNS biology that can complement CSF and PET in Alzheimer's disease research.
- Mechanistic insight comes from treating EV cargo as a system (ATN plus neuroinflammation), not as isolated Aβ or tau measurements.
- Synaptic proteins in EVs are increasingly valuable because synaptic dysfunction tracks cognitive decline more closely than plaque burden.
- The main technical challenge is not mass spectrometry alone, but upstream specificity: enriching rare CNS-derived EVs from plasma while controlling for soluble antigens and lipoprotein-associated background.
- A pragmatic translational workflow is discovery LC–MS/MS → orthogonal CSF correlation → targeted PRM verification in large cohorts.
The Blood-Brain Barrier (BBB) and the Liquid Biopsy Paradigm (brain-derived exosomes in Alzheimer's disease)
The major analytical roadblock in neurodegenerative disease research isn't a lack of biomarkers. It's access.
The blood-brain barrier (BBB) is built to isolate the brain from peripheral fluctuations in proteins, lipids, and inflammatory mediators. That protection is great for physiology, but it creates a practical bottleneck for translational studies: if you want to measure CNS biochemistry directly, you typically rely on cerebrospinal fluid (CSF) collection or positron emission tomography (PET). Both approaches have clear value, and both come with real friction in preclinical and longitudinal human cohorts.
Brain-derived exosomes (BDEs) and neuron-derived extracellular vesicles (NDEVs) create a different window for a blood-based liquid biopsy for Alzheimer's disease. Small extracellular vesicles can traverse the BBB bidirectionally and enter peripheral blood while retaining a membrane-protected cargo of proteins and post-translational modifications. In biomarker terms, that means a venous blood draw can contain a low-abundance, but information-rich, proteomic snapshot of what stressed neurons and glia are packaging and releasing.
For large-scale preclinical cohorts, that changes what's feasible. Serial blood draws allow longitudinal tracking of brain pathology at timepoints where CSF collection would be burdensome and imaging would be cost-prohibitive. The translational value isn't only convenience. It's temporal resolution: more sampling points makes it easier to catch early inflection points, treatment responses, and divergence between animals that would otherwise look similar.
Key Takeaway: BDE/NDEV analysis is valuable because it enables brain-associated molecular processes to be monitored longitudinally through blood-based samples, provided that enrichment and QC are strict enough to preserve CNS specificity.
Decoding the CNS Exosomal Cargo: Beyond Amyloid and Tau
Aβ and tau are foundational hallmarks of Alzheimer's disease (AD), but a single-marker mindset can hide what actually drives functional decline. A more useful framing is the ATN model (Amyloid/Tau/Neurodegeneration), expanded in practice to include neuroinflammation. In EV proteomics, this naturally becomes a multi-axis cargo readout: amyloid peptides and APP fragments, tau phosphorylation patterns, synaptic integrity markers, lysosomal stress proteins, and glia-derived inflammatory signals.
The advantage of EVs here is compositional, which is why exosome cargo proteomics for Alzheimer's has become a practical discovery strategy. You don't have to choose one axis early. With the right enrichment and mass spectrometry strategy, you can map multiple cargo categories from the same biological draw, then decide which panel has the best mechanistic and translational behavior.
Aβ and the "Prion-Like" Propagation of p-Tau
Exosomes can begin as a neuronal coping strategy. Neurons and glia use vesicle trafficking to move proteins out of the cytosol, including aggregation-prone species. Early on, packaging toxic Aβ fragments into EVs can be understood as a clearance-associated route.
However, EV-mediated packaging and release may also support pathological protein organization and intercellular spread. Classic work showed that Aβ peptides can be released in association with exosomes, placing aggregation-prone material into an extracellular environment where it can persist and interact with recipient cells and extracellular matrices (Rajendran et al., PNAS, 2006). In experimental systems, exosomal membranes can provide a scaffold that concentrates Aβ and supports nucleation, helping explain why EV-associated Aβ may contribute to plaque dynamics.
Tau adds a second propagation layer. The "prion-like" spreading hypothesis proposes that misfolded tau seeds transmit along connected neural networks, driving regional progression. Exosomes are plausible carriers because they can package toxic tau species and deliver them to new cells. Evidence for exosomal tau and disease-relevant phosphorylation in early AD includes tauopathy model work reported by Saman et al. (J Biol Chem, 2012).
For translational proteomics, the highest-value question is rarely "is tau present?" It's "which tau species, and which phosphorylation sites?" Early- and late-stage phosphorylation patterns can provide a staging-like dimension. For example:
- p-Tau181 is widely studied as an AD-relevant phospho-epitope in biofluids.
- p-Tau217 is strongly validated as a blood biomarker in many contexts, and is increasingly assessed in EV-associated pools.
- p-Tau396 has been reported in EV biomarker studies and is often discussed as a later-stage phosphorylation context.
Mass spectrometry enables site-specific resolution that antibody assays typically can't match, especially when multiple phospho-sites and nearby modifications co-exist on the same peptide backbone.
Synaptic Proteins: The Correlates of Cognitive Decline
If you want a marker that follows cognition, synapses are hard to ignore.
Aβ and tau loads can plateau or decouple from short-term functional change, especially once pathology is established. Synaptic dysfunction, in contrast, tracks the clinical trajectory: the loss of synaptic integrity aligns closely with cognitive impairment. That's why synaptic proteins in EVs are becoming high-priority targets for biomarker panels.
From a proteomics perspective, the appeal is practical as well as biological. Synaptic proteins can be quantified as a multiplexed panel, and shifts in a panel can be more informative than a single marker that saturates.
Key synaptic cargo detectable via proteomics includes:
- Presynaptic markers: SNAP-25, GAP-43, SV2A
- Postsynaptic markers: neurogranin, GluR2 (AMPA receptor subunit)
A particularly interesting case is SV2A. In vivo, synaptic PET imaging using SV2A ligands has emerged as a way to estimate synaptic density, but PET is expensive and difficult to scale. Detecting SV2A in EV-enriched fractions opens a path toward a blood-accessible proxy, with the important caveat that analytical specificity and validation determine whether the signal is truly CNS-derived.
A practical way to use synaptic cargo in study design is to treat it as a "dynamic axis" alongside ATN. Aβ/tau can anchor disease biology; synaptic panels can anchor functional relevance.
| Cargo axis | Examples in EV proteomics | What it helps you answer | Common failure mode |
|---|---|---|---|
| Amyloid | Aβ species, APP fragments | Is amyloidogenic biology present or changing? | Over-interpreting a small signal without confirming EV purity |
| Tau phosphorylation | p-Tau181, p-Tau217, p-Tau396 site-level peptides | Is tauopathy biology progressing, and in what pattern? | Missing low-abundance phosphopeptides without optimized enrichment |
| Synaptic integrity | SV2A, SNAP-25, GAP-43, neurogranin | Is the biology tracking functional decline or treatment response? | Treating single proteins as definitive without panel behavior |
| Neuroinflammation | glial EV markers, cytokine-associated proteins | Is neuroimmune activation changing over time? | Conflating peripheral inflammation with CNS immune responses |
Astrocytic and Microglial EVs: The Neuroinflammation Window
The EV story in AD isn't neuron-only.
Astrocytes and microglia actively shape synaptic homeostasis, clearance pathways, and inflammatory signaling. Their extracellular vesicles can carry immune- and stress-related proteins that complement ATN readouts. In practice, glial-derived EV analysis allows investigators to monitor neuroinflammation non-invasively, which matters because neuroimmune modulation is a major focus in AD drug development.
This adds an important design choice. If you restrict yourself to neuron-enriched fractions, you may miss the inflammatory axis. If you enrich multiple cell-derived EV populations (or capture different fractions in parallel), you can ask whether treatment shifts immune activation independently of amyloid and tau.
The Analytical Bottleneck: Isolating CNS-Derived EVs from Plasma
The decisive technical challenge is abundance. CNS-derived EVs are a tiny fraction of total circulating vesicles in plasma. Most of what you capture from blood is not brain-derived unless your enrichment is rigorous.
That's why EV biomarker programs live or die on upstream strategy: reducing plasma background, capturing a defined subpopulation, and proving you actually captured what you think you captured.
In service terms, the workflow often starts with clean isolation and purification steps that reduce soluble proteins and lipoprotein background before enrichment. For teams building CNS-derived EV studies, the logic of method selection (SEC vs gradient vs immunocapture) matters at least as much as the downstream MS platform. This is where Exosome Isolation and Purification Service can fit into a broader discovery workflow: Exosome Isolation and Purification Service.
Navigating the L1CAM Controversy
L1CAM (CD171) has been widely used as an immunocapture target for neuronal EVs, largely because it is neuron-associated and has accessible epitopes.
The controversy is not about whether L1CAM exists in blood. It's about what form it takes. A central critique is that a substantial fraction of circulating L1CAM signal may come from soluble cleavage fragments rather than vesicle-bound protein, which can cause immunocapture to pull down non-EV material.
For biomarker teams, this isn't an academic argument. It changes how you design capture and controls.
CRO-level mitigation strategies that increase specificity include:
- Dual-marker capture: pair L1CAM with a second neuronal transmembrane marker to reduce soluble-antigen artifacts.
- Alternative neuronal markers: NCAM and receptor-linked markers (for example GluR2 in certain contexts) can be used as orthogonal capture targets.
- Fraction-aware QC: confirm that your antigen and EV markers co-elute in EV-containing fractions rather than protein-rich fractions.
The key operational issue is not identifying a universally superior marker, but validating marker performance for each biofluid, cohort, and capture format. It's that you treat marker choice as a hypothesis that must be validated per biofluid, cohort, and capture format.
Validation via Untargeted Proteomics
If the goal is to establish a defensible NDEV/BDEV proteomic profile, ELISA alone is usually insufficient.
A strong ELISA signal can tell you the capture antibody bound something, but it can't tell you whether:
- the isolate has a CNS-like proteomic signature,
- the fraction is dominated by plasma proteins,
- lipoproteins or abundant blood proteins are driving apparent differences,
- the capture is biased toward a subpopulation that changes with disease unrelated to AD biology.
Untargeted LC–MS/MS can answer those questions because it reads out the whole proteome of the fraction. Practically, untargeted proteomics gives you:
- positive evidence (neuronal and EV marker enrichment)
- negative evidence (absence or reduction of plasma contamination signatures)
- an unbiased starting point for panel selection
This is also where high-quality EV characterization becomes a backbone step. Teams commonly combine orthogonal characterization and discovery proteomics so they can defend both "what we isolated" and "what we found." If you're planning a combined characterization + multi-omics workflow, Exosome Analysis Services provides the upstream characterization components that help keep discovery interpretable: Exosome Analysis Services.
Translational Workflows: From Discovery to PRM Verification
A translational EV biomarker program has a familiar arc:
- discovery and mechanistic mapping
- verification in larger cohorts
- orthogonal correlation to prove biological relevance
EVs don't change this arc. They raise the bar on upstream specificity and analytical sensitivity.
CSF-to-Plasma Orthogonal Correlation
The gold-standard design is simple in principle: if you discover EV cargo changes in plasma, correlate them with matched CSF samples from the same cohort.
This does two things.
First, it reduces the risk that your plasma EV signal is driven by peripheral biology. Second, it turns a plausible biomarker into a CNS-relevant biomarker by showing that the direction and magnitude of change align with CNS-facing readouts.
In practice, orthogonal correlation often benefits from a structured panel: selected tau phosphopeptides, synaptic proteins, and a small set of neuroinflammation markers, rather than one molecule.
Targeted MS (PRM) for Large-Cohort Verification
Discovery proteomics identifies candidate cargo features, while verification determines whether those candidates can be quantified reproducibly across larger sample sets.
Parallel reaction monitoring (PRM) mass spectrometry is widely used for that purpose because it provides targeted, high-specificity quantification at low abundance. For tau phosphorylation-site questions, PRM can be particularly valuable: it can quantify specific modified peptides in a way that avoids many antibody cross-reactivity and multiplexing limits.
Operationally, PRM succeeds when upstream enrichment and sample preparation are tuned for low-input EV material: lysis conditions that solubilize membrane-associated proteins, digestion strategies that preserve phosphopeptides, and acquisition parameters that keep sensitivity high.
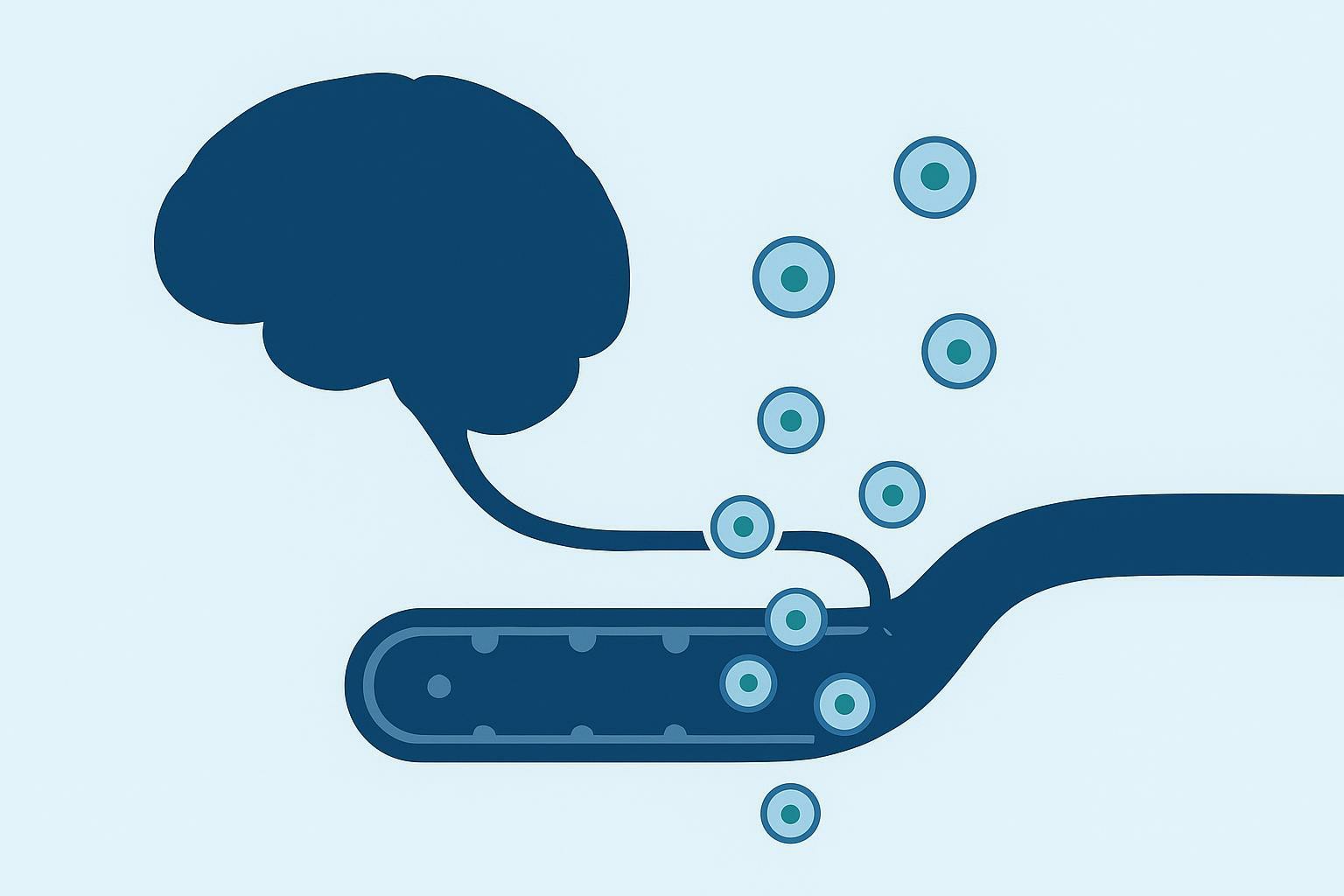
A practical way to map the end-to-end workflow is:
| Stage | What you do | What you get | What to document |
|---|---|---|---|
| Pre-cleanup | Reduce free plasma proteins/lipoproteins (e.g., SEC) | Cleaner input for capture | Fractions used and rationale |
| Enrichment | Immunocapture with single/dual markers; controls | Defined EV subpopulation | Marker rationale + isotype controls |
| Discovery | Untargeted LC–MS/MS | Candidate cargo list + contamination profile | Proteome signatures and negative markers |
| Orthogonal check | Match to CSF (same cohort) | CNS relevance confidence | Correlation strategy and caveats |
| Verification | PRM on selected peptides/panels | Quant-ready panel behavior | Calibration/QC plan and acceptance criteria |
If you're planning to combine EV enrichment with discovery proteomics and PRM verification under one workflow, Extracellular Vesicles Proteomics Services is the most direct internal resource to align expectations around LC–MS/MS and targeted phosphoproteomics: Extracellular Vesicles Proteomics Services.
Pro Tip: Treat "NDEV purity" as a quantitative QC item, not a label. Build a proteomics-based signature check (positive neuronal/EV markers plus negative plasma/lipoprotein signatures) into the workflow early, before you scale a cohort.
Workflow Support for AD-Focused EV Proteomics
Mapping the AD-associated exosomal proteome at useful depth requires highly specific EV enrichment, careful contamination control, and sensitive LC–MS/MS or targeted MS workflows. For preclinical cohort studies or translational verification panels, working with an experienced EV proteomics team can help standardize enrichment, QC, and downstream data generation.
For AD-focused EV programs, Creative Proteomics' Alzheimer's Disease Exosome Research Solution. can help align service components with study design requirements.
Frequently Asked Questions
Why study synaptic proteins in exosomes instead of just measuring Aβ and Tau?
Synaptic proteins in EVs often remain dynamic when amyloid and tau signals plateau, and synaptic dysfunction aligns more tightly with cognitive decline. Practically, a synaptic panel (for example SNAP-25, GAP-43, SV2A, neurogranin) can improve longitudinal tracking of functional trajectory, especially when used alongside amyloid- and tau-related readouts.
Is L1CAM still a reliable marker for isolating neuron-derived exosomes from blood?
L1CAM can enrich neuronal EV subpopulations in some workflows, but it is not a universal "safe" marker because soluble L1CAM forms in biofluids can confound capture. For proteomics-grade specificity, use orthogonal validation (fractionation-aware QC and proteomics signatures) and consider dual-marker capture or alternative neuronal markers such as NCAM when appropriate.
Can mass spectrometry detect specific phosphorylation sites of Tau in circulating exosomes?
Yes. Site-specific tau phosphopeptides can be detected and quantified, but success depends on enrichment specificity and MS sensitivity. A common translational approach is untargeted LC–MS/MS for discovery, followed by targeted PRM for low-abundance phosphorylation-site verification across larger cohorts.
Do brain-derived exosomes actually cross the blood-brain barrier, or are blood EVs mostly peripheral?
EVs can cross the BBB, and CNS-derived EVs have been detected in blood. The practical issue is abundance: CNS-derived EVs are rare in plasma, so enrichment and validation determine whether your signal is genuinely CNS-linked.
What is the biggest reason NDEV biomarkers fail to replicate across studies?
Method variability. Differences in plasma handling, pre-cleanup, capture markers, and how "purity" is defined can change the apparent biomarker signal. Using proteomics-based validation and standardized controls helps reduce between-study drift.
Should an AD EV biomarker study focus on neurons only, or include glial EVs too?
Include glial EVs when neuroinflammation is part of the biological hypothesis or the drug mechanism of action. Microglia- and astrocyte-derived EV cargo can add an immune-response axis that complements ATN and synaptic panels.
How do you prove your captured fraction is truly CNS-enriched?
Use orthogonal evidence. Combine EV characterization (particle sizing and morphology), marker validation with appropriate controls, and untargeted LC–MS/MS to show a CNS-relevant proteome signature with minimal plasma/lipoprotein contamination.
What's a reasonable marker strategy when you're worried about soluble antigen contamination?
Use redundancy. Pair a primary capture marker with an orthogonal neuronal transmembrane marker, and validate with proteomics signatures rather than relying on a single immunoassay readout.
References
- Alzheimer's disease beta-amyloid peptides are released in association with exosomes
- Exosome-associated tau is secreted in tauopathy models and is selectively phosphorylated in CSF of early Alzheimer's disease
- Depletion of microglia and inhibition of exosome synthesis halt tau propagation
- Brain-Derived Exosomal Proteins as Effective Biomarkers for Diagnosing Alzheimer's Disease
- Emerging blood exosome-based biomarkers for preclinical and clinical Alzheimer's disease: a systematic review and meta-analysis
- Most L1CAM Is not Associated with Extracellular Vesicles in Human Biofluids
- Examination of the enrichment of neuronal extracellular vesicles using NCAM
- Increased levels of the synaptic proteins PSD-95, SNAP-25, and neurogranin in Alzheimer's disease
* For Research Use Only. Not for use in diagnostic procedures.
