Waxes are a chemically diverse class of lipids defined not by a common molecular scaffold but by a shared set of physical properties: extreme hydrophobicity, solidity at ambient temperature, and resistance to chemical and enzymatic degradation. These properties derive from their molecular architecture — long-chain fatty acids esterified to long-chain fatty alcohols, producing molecules with two extended hydrocarbon chains connected by a single ester bond. The resulting structures contain 40-64 carbon atoms and are among the most hydrophobic molecules produced by living organisms. The existing resource on this topic provides a foundational overview of wax classification, chemical composition, and biological function. This guide extends that foundation by examining the biosynthetic origin of wax components through the very long-chain fatty acid (VLCFA) elongation pathway, by comparing the structural and metabolic logic of waxes with that of triglycerides, and by discussing how lipidomics approaches — particularly LC-MS/MS — enable the characterization of wax composition in biological samples. All methods and analyses described are for research use only.
Lipidomics profiling services provide comprehensive wax and neutral lipid analysis using validated LC-MS/MS methods for plant, animal, and microbial research samples.
Figure 1: Very long-chain fatty acid biosynthesis — from plastid FAS to ER fatty acid elongase complex

What is Wax — A Concise Definition and Classification
The existing resource provides detailed coverage of wax classification systems. For reference, waxes are divided into three broad categories: natural waxes (plant cuticular waxes, animal waxes such as beeswax and lanolin, and mineral waxes derived from petroleum), microbial waxes (produced by certain bacteria and yeasts under specific growth conditions), and synthetic waxes (polyethylene waxes, Fischer-Tropsch waxes, and microcrystalline waxes manufactured for industrial applications). This guide does not repeat the classification details. Instead, it focuses on the biochemical logic that unifies these diverse materials: all waxes derive their properties from very long-chain fatty acids and their derivatives, and the specific composition of VLCFA chain lengths and functional groups in a wax determines its physical properties, biological function, and analytical behavior.
A single plant cuticular wax is not a single molecular species but a complex mixture of 50-100 different compounds, including n-alkanes (typically C25-C35, comprising 30-50% of the total wax), aldehydes, primary alcohols, secondary alcohols, ketones, wax esters (C40-C54), and triterpenoids. The relative proportions of these compound classes vary by species, tissue, developmental stage, and environmental conditions. In Arabidopsis thaliana leaf wax, alkanes and primary alcohols predominate; in maize silk wax, very long-chain fatty acids and aldehydes are the major components. This compositional complexity is the direct result of the biosynthetic pathways that generate VLCFA derivatives, and understanding these pathways is essential for interpreting wax composition data from lipidomics experiments.
Very Long-Chain Fatty Acids — The Biosynthetic Building Blocks of Waxes
The defining chemical feature of wax components is their derivation from very long-chain fatty acids — saturated fatty acids with chain lengths of 20 to 34 carbon atoms. These VLCFAs are not produced by the same fatty acid synthase (FAS) complex that synthesizes the standard C16 and C18 fatty acids used for membrane lipids and triacylglycerols. Instead, VLCFAs are generated by a dedicated fatty acid elongation system located in the endoplasmic reticulum that extends pre-existing C16 and C18 fatty acyl-CoAs by sequential addition of two-carbon units.
The C16:0 and C18:1 fatty acids that serve as the precursors for VLCFA synthesis are produced by the type II fatty acid synthase complex in the plastid through the standard acetyl-CoA carboxylase and fatty acid synthase reactions. These fatty acids are exported from the plastid as acyl-CoA thioesters and enter the endoplasmic reticulum, where the fatty acid elongase (FAE) complex catalyzes the extension of the acyl chain. Each elongation cycle adds two carbon atoms to the growing chain and requires four sequential enzymatic activities: 3-ketoacyl-CoA synthase (KCS), which condenses malonyl-CoA with the growing acyl-CoA to form a β-ketoacyl-CoA intermediate; 3-ketoacyl-CoA reductase (KCR), which reduces the ketone to a hydroxyl group; 3-hydroxyacyl-CoA dehydatase (HCD), which removes water to form an enoyl-CoA; and enoyl-CoA reductase (ECR), which reduces the double bond to form the fully saturated fatty acyl-CoA that is two carbons longer than the substrate.
The KCS enzyme is the rate-limiting and chain-length-determining step of the elongation cycle. Different KCS isoforms have different substrate specificities for acyl-CoA chain length, and the complement of KCS genes expressed in a given cell type determines the VLCFA chain length distribution. Arabidopsis thaliana encodes 21 KCS genes, each responsible for producing VLCFAs of a specific chain length range — KCS1 produces C22-C24, KCS6/CER6 produces C26-C28 for cuticular wax biosynthesis, and KCS10/FDH is required for C22-C24 VLCFAs in sphingolipid biosynthesis. A 2024 study in Plant, Cell and Environment demonstrated that KCS19 contributes to C24-C28 VLCFA production in cotton cuticular wax, and that overexpression of KCS19 increases total cuticular wax load by approximately 30%. A 2025 study in Plant Science further showed that the VAP27-1 protein interacts with KCS6 and CER2 (an acyltransferase that modifies the KCS substrate) to facilitate VLCFA biosynthesis for cuticular wax production in Arabidopsis, providing evidence for a protein complex that organizes the spatial clustering of FAE components on the ER membrane.
The chain length of the VLCFA products directly determines the physical properties of the waxes derived from them. Longer chains produce waxes with higher melting points and greater hydrophobicity because the increased number of van der Waals interactions between adjacent hydrocarbon chains strengthens the crystalline packing of the wax layer. A wax composed predominantly of C28-C30 alkanes has a melting point of approximately 60-65°C and forms a relatively permeable barrier. A wax composed of C32-C34 alkanes has a melting point of approximately 75-85°C and forms a substantially more effective water barrier. The chain length distribution is environmentally regulated — drought stress upregulates specific KCS isoforms that produce longer-chain VLCFAs, increasing both the total wax load and the average chain length, which collectively reduce cuticular water loss by 30-50% in drought-adapted plants. Lipidomics analysis of VLCFA composition can detect these environmentally induced chain length shifts as an indicator of plant stress adaptation.
Figure 2: Plant cuticular wax biosynthesis — spatial organization from plastid through ER to cuticle deposition
Waxes vs Triglycerides — Structural, Functional, and Metabolic Contrasts
Waxes and triglycerides (triacylglycerols, TAGs) are both composed of fatty acids esterified to an alcohol, but the identity of the alcohol determines their fundamentally different biological roles. In TAGs, three fatty acids are esterified to a single glycerol molecule, producing a relatively compact structure with a polar glycerol head group and three non-polar fatty acyl tails. In waxes, one fatty acid is esterified to one long-chain fatty alcohol, producing an extended linear structure with two long hydrocarbon chains connected by a single ester bond and no residual polar group of significance. This structural difference has profound consequences for metabolism, function, and analytical detection.
The glycerol backbone of TAGs is recognized by lipases — enzymes that hydrolyze the ester bonds to release free fatty acids for β-oxidation. Adipose triglyceride lipase (ATGL), hormone-sensitive lipase (HSL), and monoacylglycerol lipase (MGL) sequentially remove fatty acids from TAGs, making TAGs the primary energy storage lipid in eukaryotes. The wax ester bond, by contrast, is recognized only by wax ester hydrolases, a much smaller enzyme family with limited tissue distribution. Most organisms lack significant wax ester hydrolase activity, which means that waxes are metabolically inert — they cannot be mobilized for energy production even under starvation conditions. This metabolic stability is the evolutionary basis for the functional specialization of waxes as structural and protective lipids rather than storage lipids. If a seed accumulated wax esters instead of TAGs for energy storage, the embryo would starve during germination because it could not access the fatty acids in the wax.
From an analytical perspective, TAGs and waxes behave differently in mass spectrometry. TAGs are readily detected in positive ion mode as ammonium adducts [M+NH₄]⁺, and collision-induced dissociation (CID) produces characteristic diglyceride fragment ions from the neutral loss of a single fatty acid, enabling the determination of the fatty acid composition. Wax esters are more challenging: they form weak ammonium adducts, and CID produces less predictable fragmentation patterns because the ester bond is located in the center of a long hydrocarbon chain rather than adjacent to a polar group. APCI (atmospheric pressure chemical ionization) is generally preferred over ESI for wax analysis because the gas-phase ionization mechanism of APCI is more efficient for the highly non-polar wax molecules. The characteristic fragments from wax ester CID are the protonated fatty acid [FA-OH₂]⁺ and the dehydrated fatty alcohol [alcohol-OH]⁺, which together enable the identification of both the acid and alcohol components of the wax ester. For the hydrocarbon components of waxes (alkanes and alkenes), APCI produces [M+H-H₂]⁺ ions that can be identified by accurate mass. Targeted lipidomics services can distinguish wax-derived and TAG-derived fatty acids in complex biological extracts through their characteristic fragmentation patterns.
Plant Cuticular Wax Biosynthesis — From Plastid to Cuticle
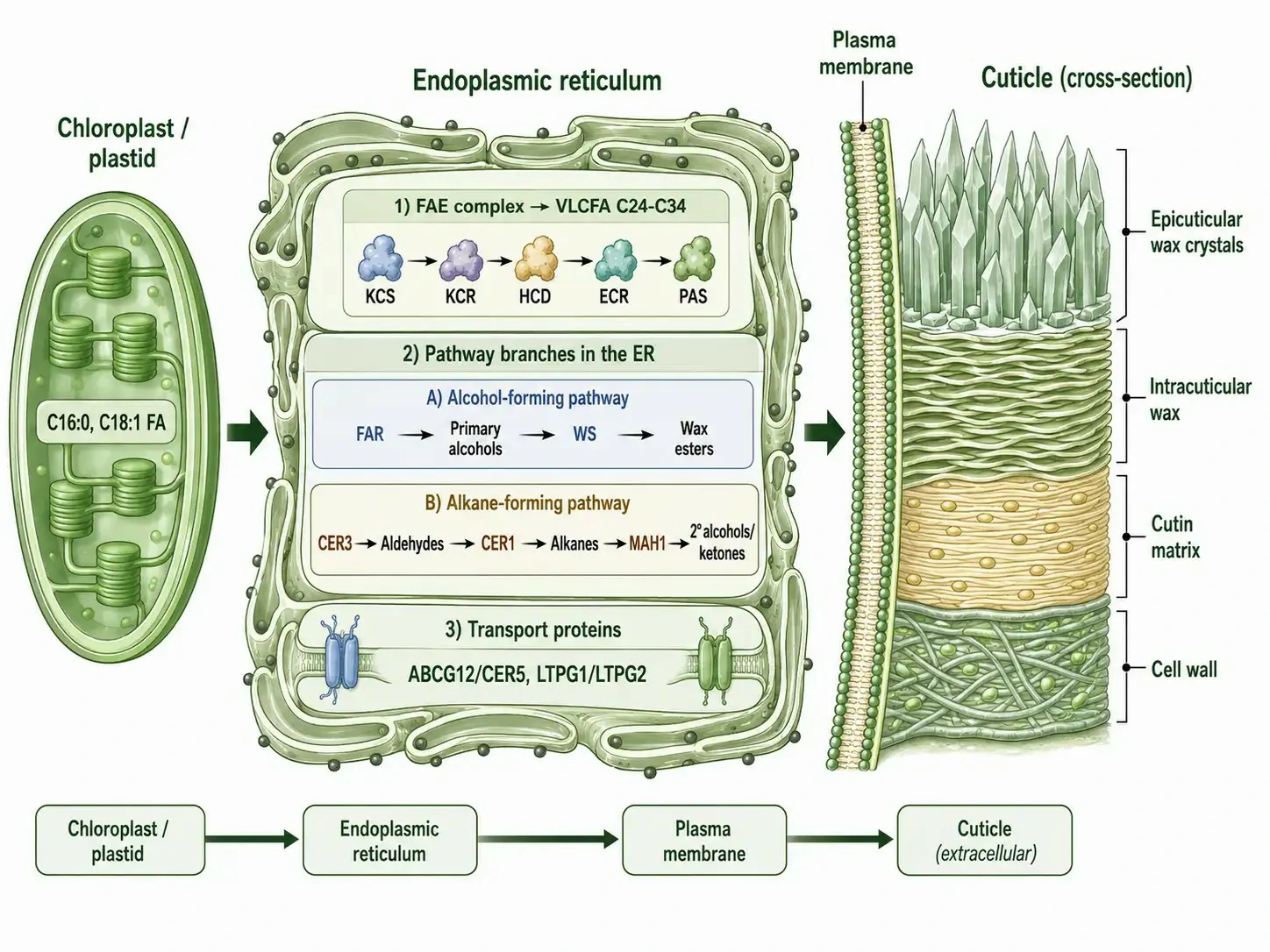
The biosynthesis of plant cuticular wax is a spatially organized process that spans three cellular compartments — the plastid, the endoplasmic reticulum, and the plasma membrane-cell wall interface — and involves at least seven distinct enzyme families. The spatial organization ensures that the highly hydrophobic wax components are synthesized in membrane environments and delivered directly to their extracellular destination without passing through the aqueous cytosol, where they would precipitate.
The biosynthetic pathway begins in the plastid, where the type II fatty acid synthase produces C16:0-ACP and C18:1-ACP. These are hydrolyzed by acyl-ACP thioesterases (FATB for saturated chains, FATA for unsaturated chains), exported from the plastid, and converted to CoA thioesters by long-chain acyl-CoA synthetases (LACS) on the outer plastid envelope. In the endoplasmic reticulum, the FAE complex extends these C16/C18 chains to C24-C34 VLCFAs, as described in the preceding section. The VLCFA-CoAs then enter one of two biosynthetic branches that generate the diverse compound classes found in cuticular wax.
The alcohol-forming pathway converts VLCFA-CoAs to primary alcohols through the action of fatty acyl-CoA reductase (FAR), an NADH-dependent enzyme that reduces the acyl-CoA to the corresponding aldehyde and then to the primary alcohol. The primary alcohols serve double duty: some are exported directly to the cuticle as wax components, while others are esterified to additional VLCFA-CoAs by wax ester synthase (WS) to form wax esters. The enzyme CER4, a fatty acyl-CoA reductase, is the major FAR responsible for cuticular wax alcohol production in Arabidopsis. In the alkane-forming pathway, VLCFA-CoAs are first reduced to aldehydes by CER3 (a bifunctional acyl-CoA reductase/aldehyde-generating enzyme), and the aldehydes are then decarbonylated to alkanes by CER1, an aldehyde decarbonylase that removes the carbonyl carbon as carbon monoxide. The released alkanes have one fewer carbon than their VLCFA precursors. Secondary alcohols and ketones are produced from alkanes by the midchain alkane hydroxylase MAH1, which inserts a hydroxyl group at specific internal positions. This secondary modification changes the physical properties of the wax layer — secondary alcohols and ketones produce a less crystalline, more amorphous wax structure that maintains flexibility at low temperatures, preventing the cracking that would occur in a purely alkane-based wax layer in freezing conditions. Plant cuticular wax analysis can quantify the relative abundance of each compound class to assess the wax biosynthetic pathway activity under different environmental conditions.
The final step in cuticular wax biosynthesis is the transport of wax components from the endoplasmic reticulum to the plant cuticle. This transport is mediated by two classes of proteins: ABC transporters of the G subfamily, particularly CER5/ABCG12, which are localized to the plasma membrane and export wax components to the extracellular space, and lipid transfer proteins such as LTPG1 and LTPG2, which are glycosylphosphatidylinositol-anchored proteins that facilitate the transfer of wax molecules across the hydrophilic cell wall to the cuticle. Mutations in CER5 reduce cuticular wax deposition by 60-80% in Arabidopsis, demonstrating that passive diffusion alone is insufficient for wax export — active transport is required to deliver these extremely hydrophobic molecules to their extracellular destination.
Figure 3: LC-MS/MS analytical workflow for wax component analysis
Analyzing Waxes by Lipidomics — LC-MS/MS Approaches
The analysis of wax composition in biological samples presents challenges that distinguish it from mainstream lipidomics. Wax components span a wide polarity range — from relatively polar free fatty acids and primary alcohols to completely non-polar alkanes — and their chromatographic behavior and ionization efficiency vary dramatically across this range.
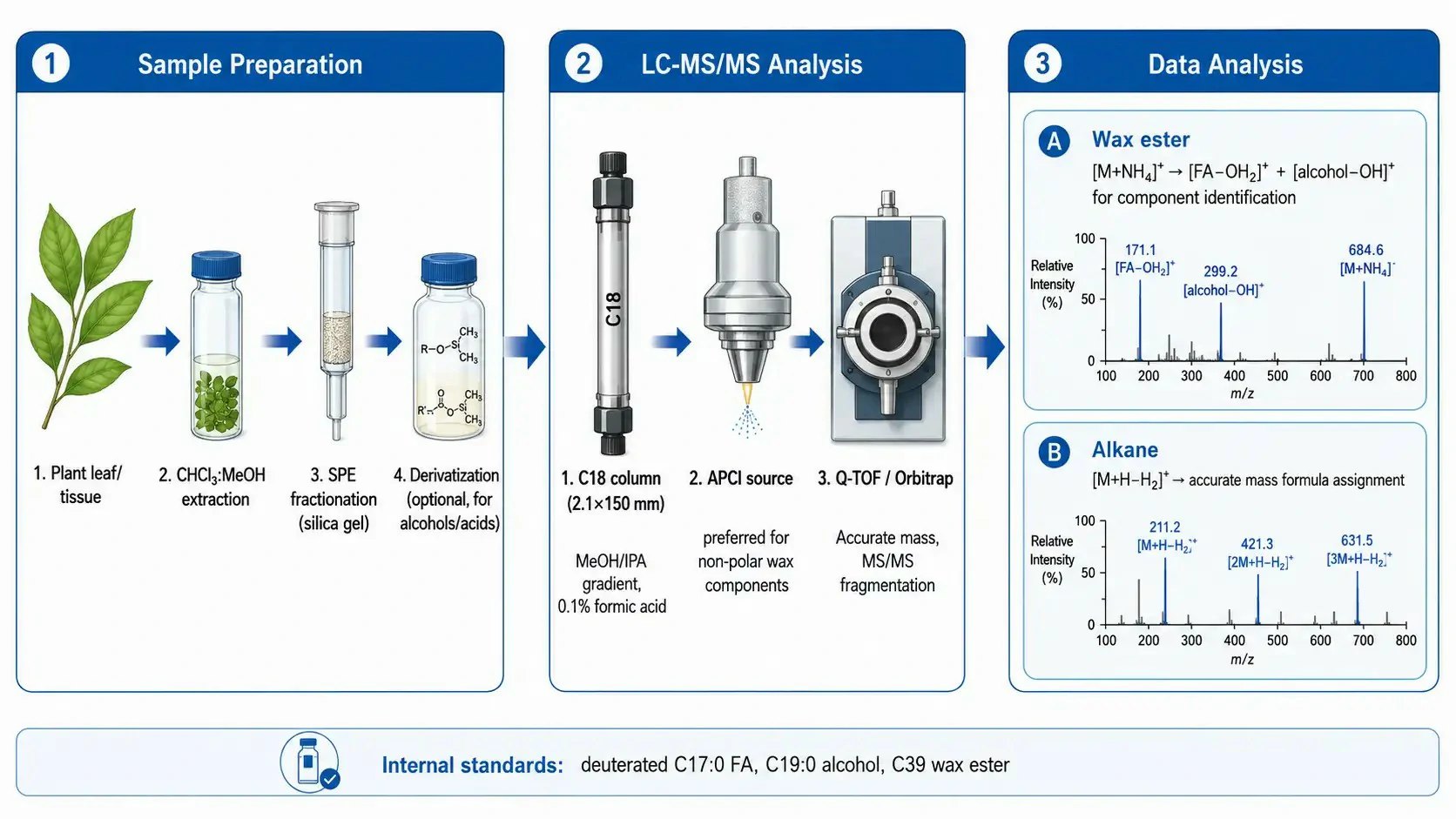
For chromatographic separation, reversed-phase liquid chromatography on a C18 column (2.1×150 mm, 1.7-2.6 µm particle size) using a methanol/isopropanol/water gradient is the standard approach. The initial mobile phase (80:20 methanol:water with 0.1% formic acid or 10 mM ammonium formate) retains the more polar wax components (free fatty acids, primary alcohols), and the gradient increases to 95:5 isopropanol:methanol over 15-20 minutes to elute the non-polar components (wax esters, alkanes). The column temperature is maintained at 40-50°C to reduce the viscosity of the organic-rich mobile phase and improve peak shape for the late-eluting non-polar compounds. For wax esters, a longer column (250 mm) or a shallower gradient may be necessary to resolve esters that differ by only two carbon atoms in chain length (e.g., C42 vs C44 wax esters), which are otherwise difficult to distinguish even by high-resolution mass spectrometry because their isotopic patterns overlap.
The choice of ionization source is critical for wax analysis. Electrospray ionization (ESI), the workhorse of lipidomics, is poorly suited for the most non-polar wax components because these molecules lack the functional groups that readily accept or donate protons in solution. Atmospheric pressure chemical ionization (APCI) is the preferred source for wax analysis because it generates ions in the gas phase through corona discharge, which can ionize non-polar molecules through charge transfer and proton transfer mechanisms that do not require the molecule to be charged in solution. In APCI positive ion mode with ammonium formate in the mobile phase, wax esters form [M+NH₄]⁺ adducts, alkanes form [M+H-H₂]⁺ ions, and fatty alcohols form [M+H-H₂O]⁺ ions. Fatty acids are detected in negative ion mode as [M-H]⁻ ions. For high-resolution instruments (Q-TOF or Orbitrap), the accurate mass of each ion can be used to assign molecular formulas, which is particularly valuable for wax analysis because the limited commercial availability of wax standards means that identification often relies on accurate mass and fragmentation patterns rather than on matching to reference spectra.
Quantitative analysis of waxes by LC-MS/MS requires internal standards that are as similar as possible to the target analytes but distinguishable by mass. Deuterated or ¹³C-labeled wax esters, alkanes, and fatty alcohols are available from specialty chemical suppliers but cover only a limited range of chain lengths. For comprehensive wax quantification, a representative set of odd-chain-length internal standards (e.g., C17:0 fatty acid, C19:0 fatty alcohol, C39 wax ester) is used, with the assumption that the ionization efficiency is comparable across chain lengths that differ by only 2-4 carbon atoms. This assumption should be validated by comparing the response factors of available even-chain and odd-chain standards of similar length. Targeted metabolomics analysis can complement lipidomics by measuring the VLCFA precursors that enter the wax biosynthetic pathway.
Figure 4: Biological functions and applications of waxes — from cuticular barrier to pharmaceutical delivery
Biological Functions and Applied Uses of Waxes
The biological functions of waxes are a direct consequence of their chemical properties: extreme hydrophobicity, metabolic stability, and the ability to form crystalline or semi-crystalline layers with specific mechanical and optical characteristics. These properties are exploited both in nature and in a range of industrial and pharmaceutical applications.
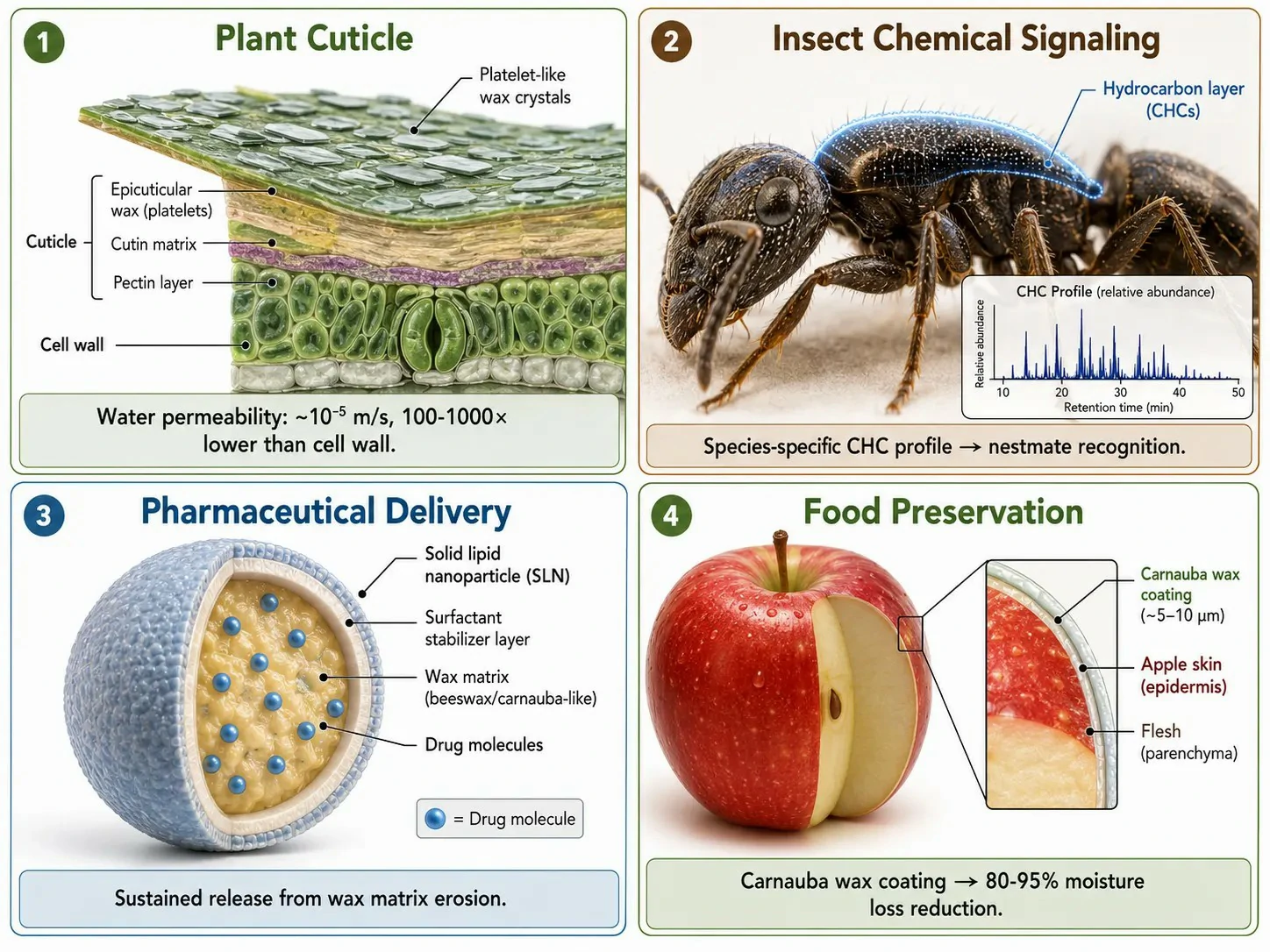
In plants, the cuticular wax layer reduces transpirational water loss by forming a hydrophobic barrier that forces water vapor to diffuse through a tortuous extracellular pathway. The water permeability of an intact plant cuticle covered with crystalline wax is approximately 10⁻⁵ m/s, which is 100-1,000 times lower than the permeability of the underlying cell wall. The effectiveness of the wax barrier depends on both the total wax load and the wax composition. Alkanes and aldehydes, which form densely packed crystalline domains, are particularly effective at reducing water permeability; triterpenoids, which disrupt crystalline packing, increase permeability but improve mechanical flexibility. The trade-off between water retention and mechanical flexibility determines the optimal wax composition for a given environment — desert plants produce waxes dominated by long-chain alkanes, while plants in temperate environments with frequent temperature fluctuations produce waxes with higher alcohol and triterpenoid content.
Animal waxes serve different but equally specialized functions. Beeswax, composed primarily of C40-C50 wax esters of palmitic acid and C26-C32 alcohols, is produced by worker bees from specialized abdominal glands and used to construct the honeycomb structure. The high melting point of beeswax (62-65°C) provides structural stability at hive temperatures (35°C), while the extreme hydrophobicity prevents water damage to the nest. Lanolin, the wax secreted by sheep sebaceous glands, coats wool fibers and provides waterproofing — the complex mixture of sterol esters, hydroxy acids, and wax diesters that constitutes lanolin has a melting point range of 38-44°C that matches the skin temperature of the sheep, ensuring that the wax remains semi-solid and flexible rather than brittle. Spermaceti, the liquid wax found in the spermaceti organ of the sperm whale, is a mixture of cetyl palmitate (C16:0 acid esterified to C16:0 alcohol) and related wax esters that remains liquid at the high pressures of deep ocean diving. The temperature-dependent density change of spermaceti was historically hypothesized to assist with buoyancy regulation, though its primary function is now understood to be acoustic — the wax-filled organ focuses and directs the sonar clicks used for echolocation.
Waxes also serve as chemical signals. The cuticular hydrocarbon profile of insects — which includes n-alkanes, methyl-branched alkanes, and alkenes — functions as a species-specific and colony-specific recognition signal in social insects such as ants, bees, and termites. These hydrocarbons are synthesized by epidermal cells, deposited on the insect cuticle, and detected by contact chemoreception. A worker ant encountering a non-nestmate detects a mismatched hydrocarbon profile and initiates an aggressive response within seconds. The chemical stability of these hydrocarbons — their resistance to degradation by environmental exposure — makes them reliable long-term signals that persist on the cuticle for the lifespan of the insect.
In applied contexts, waxes are used in pharmaceutical formulations as sustained-release matrices and as components of solid lipid nanoparticles (SLNs). SLNs are produced from biocompatible waxes such as beeswax, carnauba wax, or cetyl palmitate and can encapsulate hydrophobic drugs within their crystalline lipid matrix, releasing the drug slowly as the wax is gradually eroded or as the drug diffuses through imperfections in the crystal lattice. In food science, edible wax coatings — typically carnauba wax or shellac — are applied to fruits and vegetables to reduce moisture loss and gas exchange, extending shelf life. The wax coating reduces the respiration rate of the fruit by limiting oxygen diffusion and reduces water loss by 80-95% compared to uncoated produce. The same wax chemistry that evolved to protect plant leaves from desiccation is thus repurposed to protect harvested fruits, creating a direct connection between the fundamental biochemistry of waxes and their practical applications.
FAQ
What is the difference between a wax and a triglyceride?
Waxes are composed of one fatty acid esterified to one long-chain fatty alcohol, producing a linear structure with two hydrocarbon tails and no significant polar group. Triglycerides are three fatty acids esterified to glycerol. Waxes are metabolically inert (cannot be used for energy); triglycerides are the primary energy storage lipid.
How are very long-chain fatty acids (VLCFAs) synthesized?
VLCFAs are produced by the fatty acid elongase (FAE) complex in the endoplasmic reticulum. The FAE adds two-carbon units to C16/C18 fatty acyl-CoAs through four sequential reactions: KCS (condensation), KCR (reduction), HCD (dehydration), and ECR (reduction). Different KCS isoforms determine the final chain length.
Why can't waxes be used for energy storage like triglycerides?
Wax ester bonds are hydrolyzed only by specific wax ester hydrolases, which are absent or expressed at very low levels in most organisms. The lipases that mobilize triglycerides for energy production cannot recognize the wax ester bond, making waxes metabolically inaccessible as energy sources.
How are waxes analyzed by LC-MS/MS?
Wax components require reversed-phase C18 chromatography with methanol/isopropanol gradients and APCI ionization (rather than ESI) because of their low polarity. Wax esters produce [M+NH₄]⁺ adducts; alkanes produce [M+H-H₂]⁺ ions. High-resolution mass spectrometry enables formula assignment without requiring matched standards for every wax species.
What determines the physical properties of a wax?
The VLCFA chain length distribution and the relative abundance of different compound classes (alkanes, alcohols, wax esters, triterpenoids). Longer chains increase the melting point and hydrophobicity. Alcohols and triterpenoids disrupt crystalline packing and improve flexibility at low temperatures.
How does the plant cuticle resist water loss?
The cuticular wax layer reduces water permeability to approximately 10⁻⁵ m/s — 100-1,000 fold lower than the cell wall alone. The densely packed crystalline domains of alkanes and aldehydes force water molecules to follow a tortuous diffusion path through the wax layer.
References
- 3-ketoacyl-CoA synthase 19 contributes to cuticular wax biosynthesis in cotton. Plant, Cell and Environment. 2024;47:3764-3778.
- VAP27-1 interacts with KCS6 and CER2 to facilitate cuticular wax biosynthesis. Plant Science. 2025;354:112474.
- Dynamic relationships among pathways producing hydrocarbons and fatty acids in maize silk wax. Plant Physiology. 2024;195:2234-2250.
- Biosynthesis and secretion of plant cuticular wax. Progress in Lipid Research. 2003;42:51-80.
- Recent advances in LC-MS-based metabolomics. Mass Spectrometry Reviews. 2023;42:101-134.









